Proposal (984) to South
American Classification Committee
Note from Remsen: This is a modification for
SACC of a proposal Oscar has submitted to NACC
Treat Piaya
mexicana and P. “circe” as
separate species from P. cayana (Squirrel Cuckoo)
Description of the
problem:
A recent NACC proposal (2022-B-11;
https://americanornithology.org/wp-content/uploads/2022/03/2022-B.pdf) to split
Piaya mexicana from P. cayana failed unanimously, largely due to
a lack of genetic or vocal data, or information from the contact zone of mexicana
and thermophila. A recent paper (Sánchez‐González et al. 2023)
addressed some of these issues, and proposal 2022-B-11 overlooked genetic data
published in Smith et al. (2014). This proposal incorporates that genetic
information and additional taxonomic information from Colombia and Venezuela that
is relevant to the potential split of South American taxa. We encourage
committee members to read proposal 2022-B-11and comments on that proposal
(https://americanornithology.org/about/committees/nacc/current-prior-proposals/2022-proposals/comments-2022-b/#2022-B-11).
In particular, proposal 2022-B-11 contains photos of specimens that are
relevant to the current proposal. The introduction to this proposal includes
much of the same text as in 2022-B-11 but expands on certain topics overlooked
in 2022-B-11. Similar proposals to split Piaya cayana are being
considered concurrently by both NACC and WGAC.
![]() Piaya cayana (Linnaeus 1766) is a widespread
polytypic species found from northern Mexico to Argentina, with as many as 14
subspecies recognized (Fitzgerald et al. 2020). The species is common in
forested lowlands and foothills throughout its range. Details on relevant
subspecies are outlined here. In Middle America, the darker subspecies thermophila Sclater, 1859, is found from
eastern Mexico south to northwestern Colombia but is replaced on the Pacific
coast of the Isthmus of Tehuantepec by the pale west Mexican subspecies mexicana (Swainson, 1827), which
is found in dry forests from the Isthmus of Tehuantepec north to Sonora and
Chihuahua. Fitzgerald et al. (2020) treated thermophila as a monotypic
subspecies group, with a distribution extending south to northwestern Colombia,
where it is replaced by another monotypic subspecies group, nigricrissa
(Cabanis, 1862) of the Chocó from northwestern Colombia south to northern
Ecuador on the Pacific slope, although nigricrissa reaches as far east
as the eastern slope of the central Andes in Colombia (Chapman 1917). As the
name suggests, nigricrissa has a darker blackish vent compared to thermophila,
but it is otherwise similar. Fitzgerald et al. (2020) considered all remaining
subspecies to be part of the cayana group. In northern Colombia, thermophila
is replaced to the east by the pale rufous mehleri
Bonaparte, 1850, in the dry forests of northern Colombia and Venezuela, and
south into the Magdalena Valley of Colombia. The even paler rufous circe Bonaparte, 1850, replaces mehleri south of Lago Maracaibo. Either circe or mehleri
is found east to the Río Orinoco delta, and insulana
Hellmayr, 1906, is found on Trinidad. Subspecies mesura
(Cabanis and Heine, 1863) replaces these pale rufous taxa south across the Río
Orinoco in the northwestern Amazon Basin, likely meeting mehleri
and nigricrissa via low passes in the Andes (Chapman 1917). Compared
to nigricrissa, mesura is paler below
and has a red rather than greenish-yellow orbital skin (Ridgely and Greenfield
2001). The nominate cayana is found in the humid Guiana Shield.
Additional subspecies are found south through the remainder of South America.
Piaya cayana (Linnaeus 1766) is a widespread
polytypic species found from northern Mexico to Argentina, with as many as 14
subspecies recognized (Fitzgerald et al. 2020). The species is common in
forested lowlands and foothills throughout its range. Details on relevant
subspecies are outlined here. In Middle America, the darker subspecies thermophila Sclater, 1859, is found from
eastern Mexico south to northwestern Colombia but is replaced on the Pacific
coast of the Isthmus of Tehuantepec by the pale west Mexican subspecies mexicana (Swainson, 1827), which
is found in dry forests from the Isthmus of Tehuantepec north to Sonora and
Chihuahua. Fitzgerald et al. (2020) treated thermophila as a monotypic
subspecies group, with a distribution extending south to northwestern Colombia,
where it is replaced by another monotypic subspecies group, nigricrissa
(Cabanis, 1862) of the Chocó from northwestern Colombia south to northern
Ecuador on the Pacific slope, although nigricrissa reaches as far east
as the eastern slope of the central Andes in Colombia (Chapman 1917). As the
name suggests, nigricrissa has a darker blackish vent compared to thermophila,
but it is otherwise similar. Fitzgerald et al. (2020) considered all remaining
subspecies to be part of the cayana group. In northern Colombia, thermophila
is replaced to the east by the pale rufous mehleri
Bonaparte, 1850, in the dry forests of northern Colombia and Venezuela, and
south into the Magdalena Valley of Colombia. The even paler rufous circe Bonaparte, 1850, replaces mehleri south of Lago Maracaibo. Either circe or mehleri
is found east to the Río Orinoco delta, and insulana
Hellmayr, 1906, is found on Trinidad. Subspecies mesura
(Cabanis and Heine, 1863) replaces these pale rufous taxa south across the Río
Orinoco in the northwestern Amazon Basin, likely meeting mehleri
and nigricrissa via low passes in the Andes (Chapman 1917). Compared
to nigricrissa, mesura is paler below
and has a red rather than greenish-yellow orbital skin (Ridgely and Greenfield
2001). The nominate cayana is found in the humid Guiana Shield.
Additional subspecies are found south through the remainder of South America.
HBW-BirdLife
split mexicana from the remainder of Piaya cayana based on plumage and slight
vocal differences and parapatric distribution; citations are Navarro-Sigüenza
and Peterson (2004) and Howell (2013, in litt.): "[mexicana] differs
from parapatric subspecies thermophila
of P. cayana in its rufous underside
of tail feathers with broad black subterminal bar and broad white terminal tip
vs all-black underside of tail with broad white terminal tip (3); pale grey vs
smoky-grey lower belly and vent (2); much brighter rufous upperparts and paler
throat (1); usually greenish-grey vs greenish-yellow orbital ring (Howell 2013)
(ns1); longer tail (effect size 2.01; score 2); “somewhat different” song
(Howell 2013) (allow 1); and parapatric distribution (3)."
Piaya mexicana was described as a
species by Swainson (1827), who gave the following characters (which largely
mirror the differences described above): “Closely resembles C. cayenensis L. [=Piaya
cayana], but the tail beneath is rufous, not black; the ferruginous colour of the head and neck is likewise much brighter.”
This treatment was maintained by authors through the beginning of the 20th
century (Ridgway 1916, Cory 1919), until mexicana was lumped with P. cayana by Peters (1940). Ridgway
expanded on the differences between mexicana:
“Resembling P. cayana thermophila,
but colored portion of under surface of rectrices cinnamon-rufous (instead of
brownish black) with a dull black area immediately preceding the white tip,
general coloration much lighter, and tail relatively much longer.” Most authors
since Peters (1940) have maintained mexicana
as a subspecies of cayana.
Navarro-Sigüenza
and Peterson (2004) used Piaya cayana
as one of their case studies for contrasting a BSC classification (single
species) with a PSC/ESC classification (two species) by splitting mexicana, using this rationale:
“Populations along the Pacific lowlands from Sonora to the Isthmus of
Tehuantepec are long-tailed, pale in coloration of the underparts, whereas the
forms of eastern Mexico and Central America are shorter-tailed and darker in
color. Although a narrow contact zone is present in eastern Oaxaca between the
two forms, only one “hybrid” specimen is known and the
differences are maintained even in close parapatry.” The reference to the
“narrow” contact zone appears to be from Binford (1989), who reported a few
specimens intermediate between thermophila
and mexicana: “I have seen definite
intermediates from Rio Ostuta (MLZ 45402), Las Tejas (MLZ 54387), and Tehuantepec City (UMMZ 137345 and
137350), but some specimens from the last two localities are mexicana. Birds from Tapanatepec,
Santa Efigenia, and a point 18 mi south of Matias
Romero are close to thermophila but
very slightly paler, a condition that might represent response to the drier
environment rather than intergradation” but noted that the "abruptness and
apparent rarity of intergradation suggest that these two forms might be
separate species; a detailed study is needed.” This, combined with the
unpublished information from Howell (2013) mentioned above, appears to
constitute the basis for the HBW-BirdLife split of mexicana from the remainder of P.
cayana. NACC proposal 2022-B-11
also contains photos of two potential intermediate specimens from this region.
Ridgway
(1916) considered mexicana a species distinct from cayana, noting
that “these certainly represent two specific types; certainly it is impossible
that P. c. thermophila and P. mexicana can be conspecific, for
perfectly typical examples of each occur together in the State of Oaxaca, and
none of the large number of specimens examined shows the slightest
intergradation of characters.” In the list of specimens examined for both thermophila
and mexicana is the locality “Oaxaca; Tehuántepec”, which is where we
now know there is a limited contact zone. However, his note that there isn’t
the “slightest intergradation” does suggest that there is likely limited or no
intergradation of characters outside of this contact zone.
New information:
Sánchez‐González et al. (2023) and Smith et al.
(2014) each analyzed 1-2 mitochondrial markers from across the range of Piaya
cayana. Sánchez‐González et al.
(2023) recovered mexicana and thermophila as sister taxa, with a
divergence time of
1.24 mya (1.8 – 0.8 mya, 95% HPD), with nigricrissa unsampled. The mexicana + thermophila
clade was in turn sister to seven samples from Peru and Paraguay with a divergence time listed in the main
text of about 4.7 mya (6.5–3.2
mya, 95% HPD). However, this latter divergence time estimate appears to be an
error, based on the values shown in Figure 1. The 4.7 mya divergence date in
the figure is that of P. cayana vs. P. melanogaster, whereas the
divergence time of the Amazonian vs. the mexicana + thermophila clade
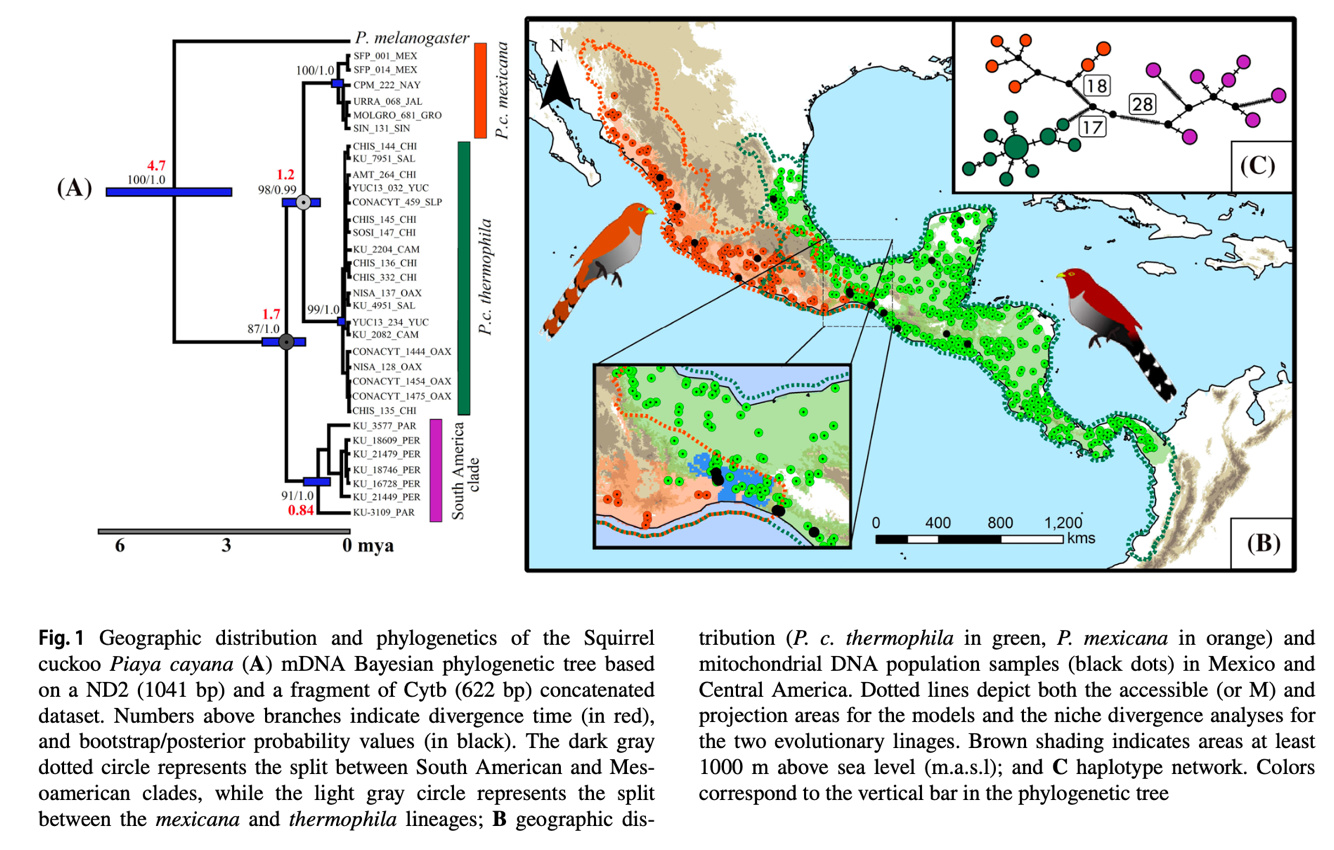
is 1.7 mya. FST and Dxy
divergence values are shown in their Table 1, and their phylogenetic tree,
haplotype network, and sampling map are shown in their Figure 1, below. The FST
results in Table 1 show FST with Nm (the number of migrants per
generation) in parentheses. However, estimates of Nm based on FST
are notoriously unreliable, especially from so few loci. See Whitlock and
McCauley (1999) for discussion of this issue.
As
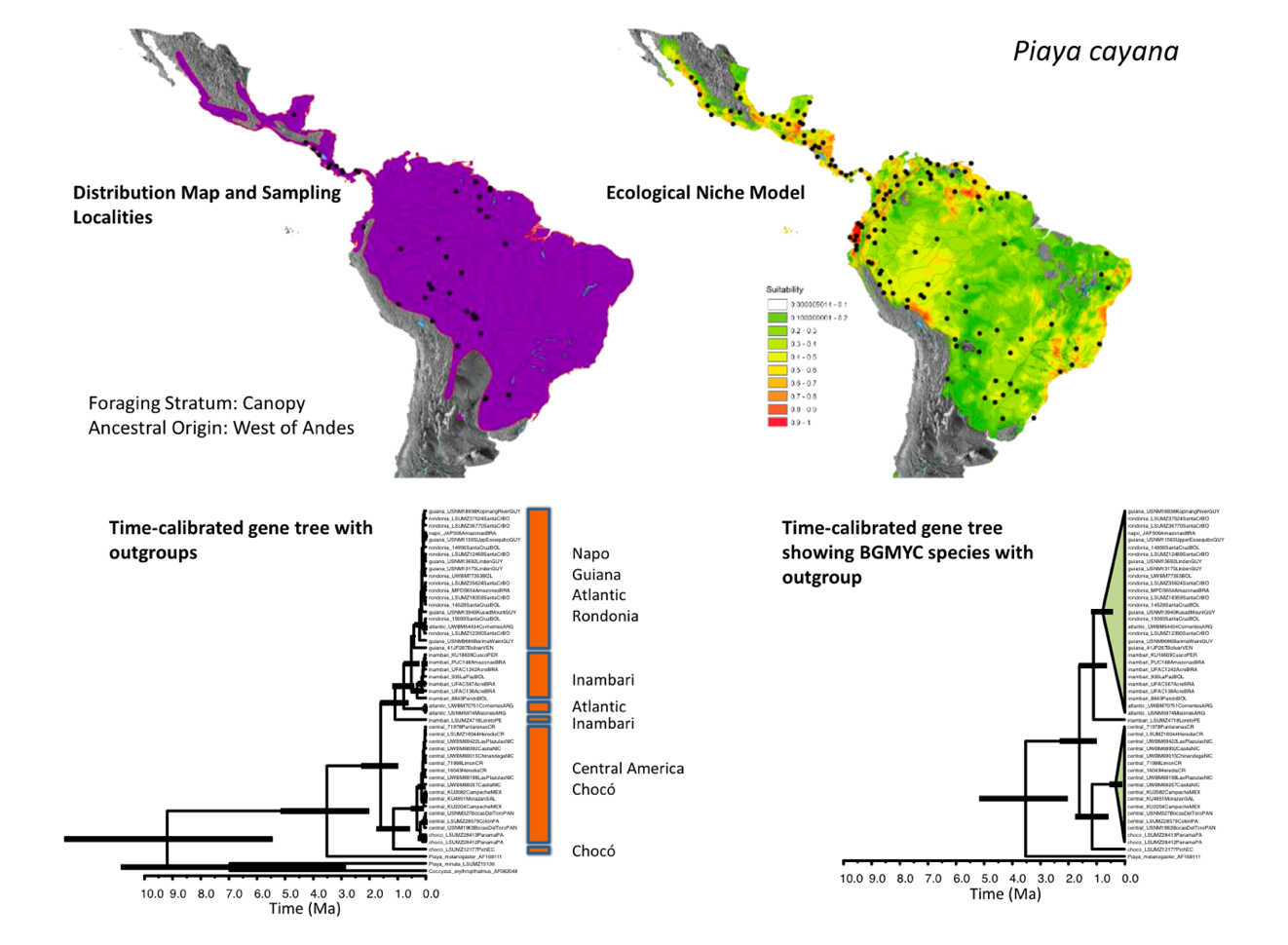
part of a broader study on tropical diversification, Smith et al. (2014)
sampled Piaya cayana from across its range, sequenced the ND2
mitochondrial gene, and used the species delimitation method bGMYC on the time-calibrated gene tree. Their results
largely agree with those of Sánchez‐González et al.
(2023), although the sampling is very different. Smith et al. (2014) sampled across much
of South and Middle America, but lacked samples from Colombia, eastern Brazil,
or western Mexico (i.e., mexicana). Smith et al. (2014) recovered four bGMYC
“species” (i.e., clades). Two of these clades contained most of their samples,
and corresponded to 1) Middle American samples (thermophila) and 2) most
of South America (much of the cayana group). The other two clades each
contained a single sample; the first was their sample from western Ecuador (nigricrissa)
which was sister to thermophila, and the second clade was a sample from
Loreto, Peru, in the northwestern Amazon. The divergence time estimates were
comparable between the two studies. These results are shown in the figure
below.

Sánchez‐González et al. (2023) also
measured specimens of thermophila and mexicana
and found significant average differences in four characters: bill width, bill
length, tail length, and the length of the white tips on the tail feathers. A
PCA of these characters largely separated the two taxa, with some overlap.
These results are shown in their Figure 2 below.
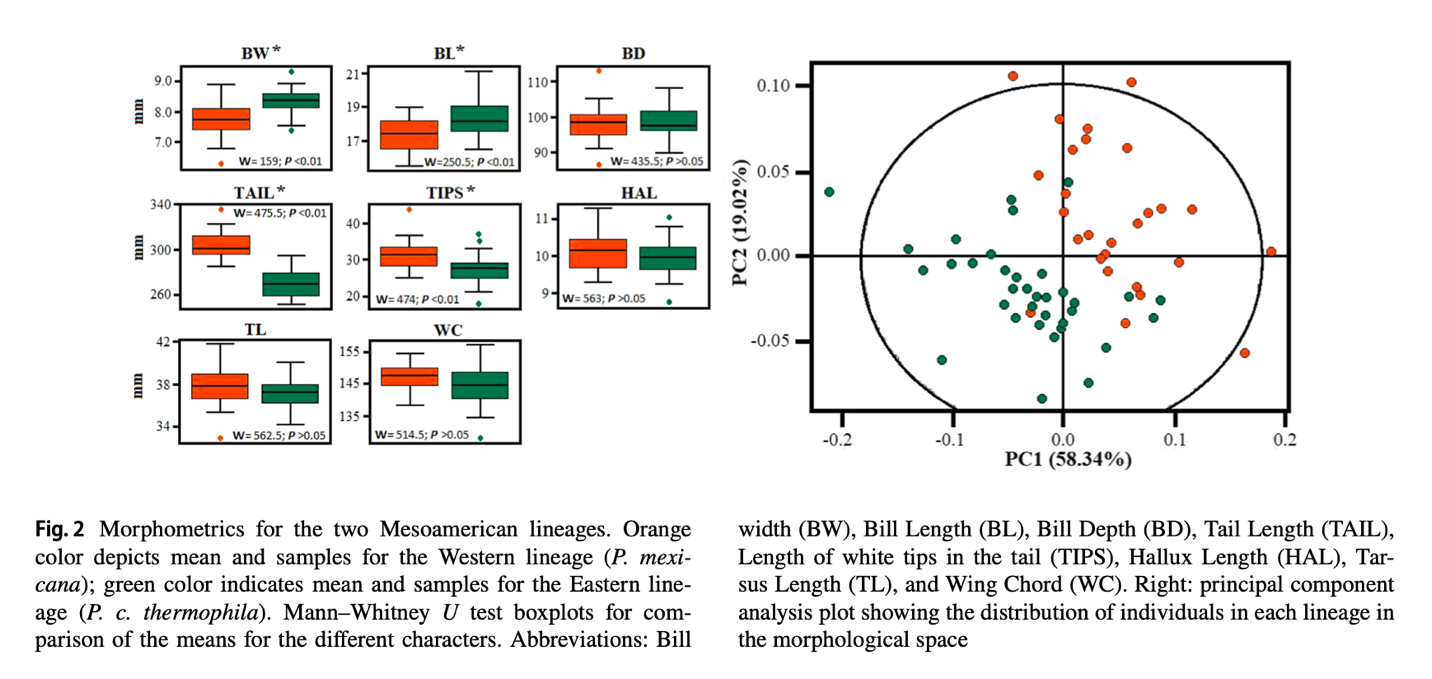
There
do not appear to be any published analyses of plumage or song from across the
distribution of P. cayana, but multiple references outlined below
discuss the plumages of each subspecies. Also, photos in 2022-B-11 nicely illustrate the plumage variation
in the group. As for song, we noted in 2022-B-11 that “the song of mexicana
appears to average higher pitched and more rapid than that of thermophila,
but some recordings of songs of thermophila seem to match recordings of mexicana”.
Pam Rasmussen in her WGAC proposal noted that mexicana “seems to have
the fastest ‘chick’ series with the sharpest (most vertical) notes, while South
American taxa seem to have more slurred (more diagonal) notes, and east Mexican
birds with longer, more resonant (less clipped) notes”, and included the following
sonograms to illustrate these differences.
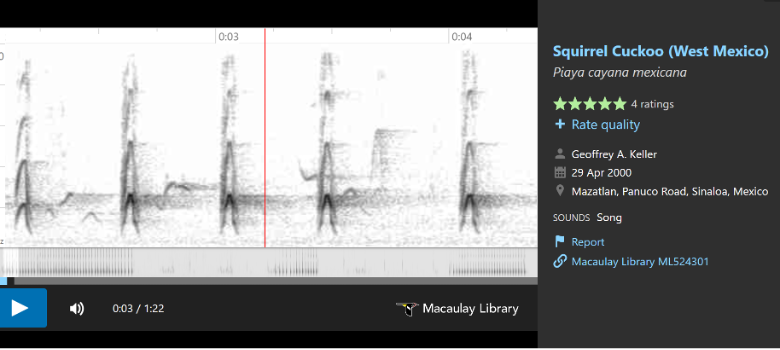
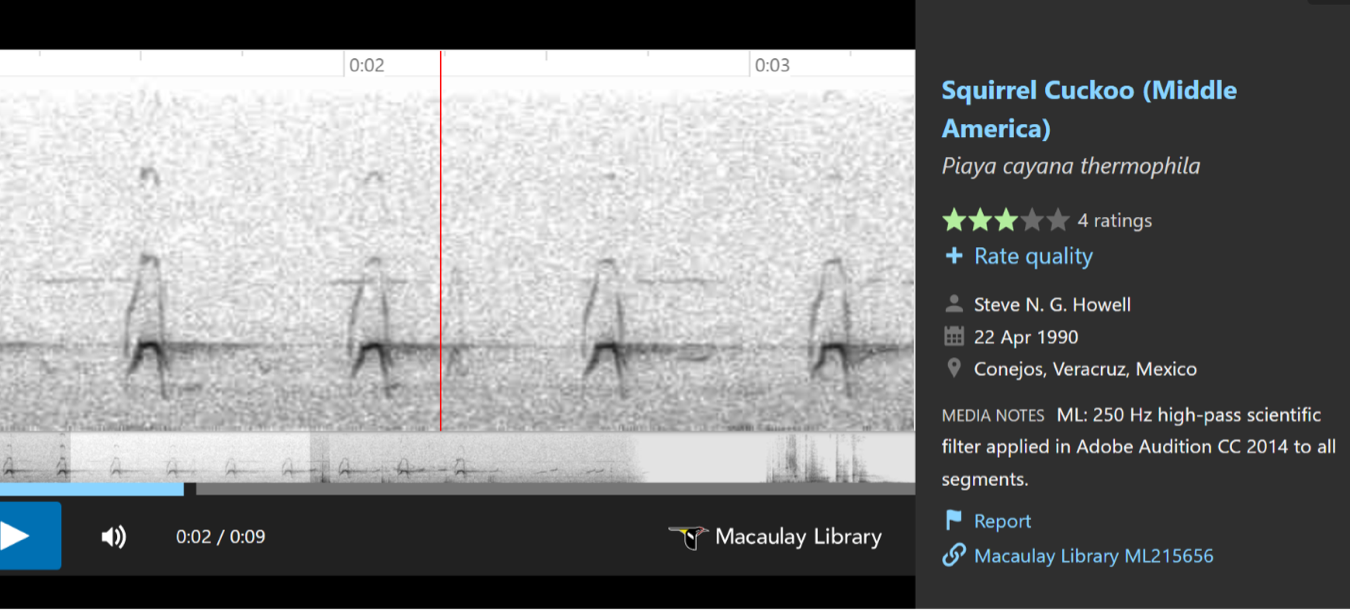
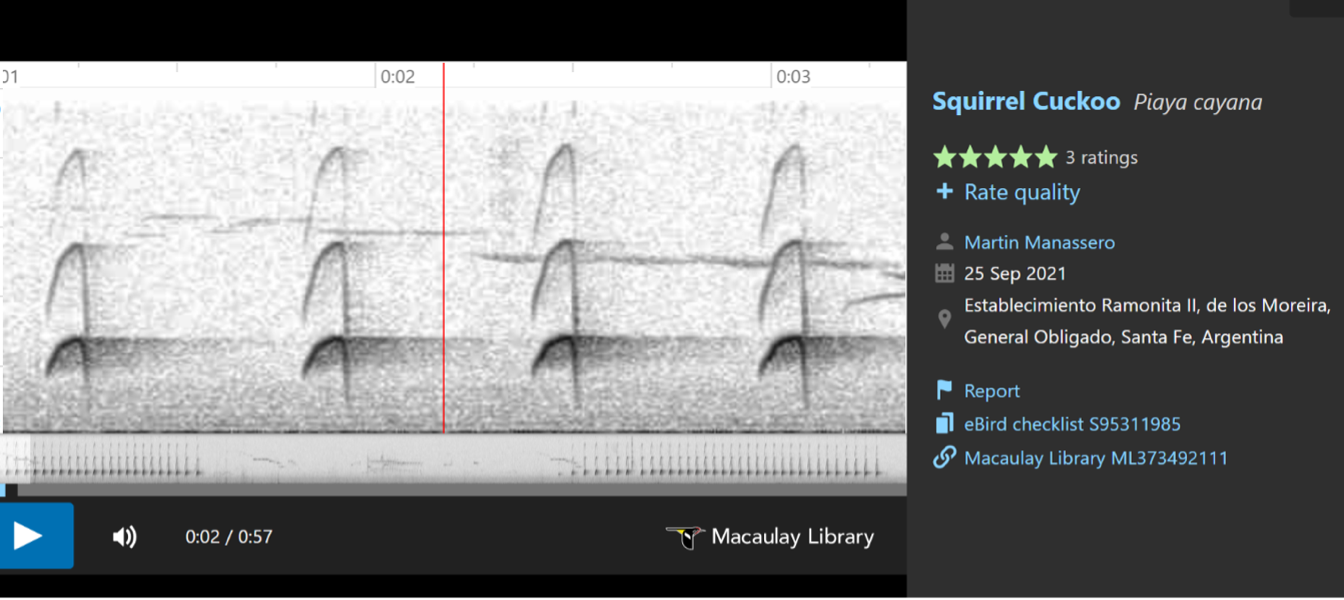
In
addition, the limited genetic data suggest that if mexicana is split,
then a split of South American taxa should also be considered, as this is a
deeper split in the mitochondrial gene tree. This split is also currently being
considered by WGAC. However, neither Smith et al. (2014) nor Sánchez‐González et al. (2023) had
samples from anywhere in Colombia, nor from the zones of contact between Middle
American and South American groups. The sole sample from Venezuela in Smith et
al. (2014) comes from south of the Río Orinoco in the far east of the country.
Because it is very relevant to the species limits and range boundaries of
groups, we here include what information is available on the distributions of
the various forms that might come into contact. Fitzgerald et al. (2020) give
the following distributional statements (and plumage differences) for the
relevant subspecies that come into contact in Colombia and Venezuela. The first
two taxa are each considered monophyletic subspecies groups by Fitzgerald et
al. (2020):
thermophila Sclater 1859; type locality Jalapa,
Veracruz, Mexico. Occurs on the Gulf and Atlantic slopes from Mexico south to
Panama and northwestern Colombia. Relatively dark rufous-chestnut above; belly
and undertail coverts dark gray to black; underside of rectrices black, white
tips to rectrices relatively narrow.
nigricrissa (Cabanis 1862); type locality Babahoyo or Esmeraldas, Ecuador. Occurs in western Colombia
(east to the slopes of the central Andes), south of northwestern Peru. Similar
to thermophila, but plumage darker; belly and undertail coverts
blackish.
cayana group:
circe Bonaparte 1850; type
locality Caracas, Venezuela. Occurs in Venezuela, south of Lake Maracaibo.
Upperparts slightly more rufous than mehleri,
but paler than nominate cayana.
mehleri Bonaparte 1850; type
locality Santa Fé de Bogota (the same type locality
as mesura?!). Occurs in northeastern Colombia,
from the Gulf of Urabá to the Magdalena valley and the west slope of the
eastern Andes, east along the coast of northern Venezuela to the Paria
Peninsula. More rufous than mexicana, with a lighter throat and breast
that grade to light gray on the belly; underside of rectrices rufous.
insulana Hellmayr 1906; type
locality Chaguaranas, Trinidad. Trinidad. Similar to cayana,
but undertail coverts black.
cayana (Linné 1766); type
locality Cayenne. Widespread, from eastern and southern Venezuela east through
the Guyanas, south to Brazil to the north bank of the
lower Amazon. Belly ashy gray; undertail coverts darker gray; colors otherwise
similar to thermophila except that the belly and undertail coverts are
not as dark; underside of rectrices black with white tips.
mesura (Cabanis and Heine
1863); type locality Bogotá, Colombia. Occurs in eastern Colombia, Ecuador, and
Peru. Similar in plumage to nigricrissa; smaller, but with overlap in
size.
Chapman
(1917) included more detail on the distribution of the Colombian taxa, and,
critically, suggested an area of potential contact between nigricrissa
and mehleri based on a fairly extensive
specimen series. Some critical passages from Chapman (1917) are below. Note
that “columbiana” is currently regarded as a synonym of mehleri.
Piaya cayana columbiana [=mehleri]
After comparison with an essentially
topotypical series from Santa Marta, I refer to this form our specimens from
the Magdalena Valley and western slope of the Eastern Andes as far south as Chicoral. These birds have the ventral region darker, the
rectrices are blacker, and a bird from Puerto Berrio
is deeper above than true columbiana. They thus show an approach toward P.
c. nigricrissa of western Colombia, which, however, is darker above and has
much more black on the ventral region.
Piaya cayana mesura
Two forms of Piaya inhabit the Bogotá
region, P. c. mesura and P. c. columbiana.
The first occurs on the eastern slopes of the Eastern Andes, and, singularly
enough, on both eastern and western slopes of the Andes at the head of the
Magdalena Valley; the second, occurs on the slopes of the Eastern Andes west of
Bogotá and in the Magdalena Valley at least as far south as Chicoral.
Piaya cayana nigricrissa
Inhabits the Tropical and Subtropical Zones in
western Ecuador and western Colombia, extending in Colombia eastward to the
eastern slope of the Central Andes. Specimens from Antioquia east of the
Western Andes approach columbiana, but on the whole, are nearer nigricrissa.
Chapman
(1917) notes that mesura is “distinguished
chiefly by the comparative blackness of all but the central tail-feathers, seen
from below, a character that at once separates it from the other Colombian
forms”. This character is apparent in the photo of mesura
in proposal 2022-B-11, especially in comparison to the specimen of nigricrissa.
This, combined with Chapman’s statement of intermediates between nigricrissa
and columbiana [=mehleri] in
Antioquia, suggests hybridization in central Colombia, likely between
populations in the Magdalena Valley (mehleri)
and the eastern slope of the central Andes (nigricrissa). As noted
above, Chapman (1917) also indicates that samples at the far southern end of
the Magdalena Valley pertain to mesura, which
crosses over the eastern Andes in this region. An additional potential contact
zone is in low passes in southern Ecuador (vicinity of Loja). It is not clear
whether there are intergrades in these areas, which do not appear to be located
at ecotones as in mexicana vs. thermophila.
Another
point, overlooked in 2022-B-11, is that mehleri
of the northern coast of Colombia (and the taxon that presumably meets thermophila
in northwestern Colombia) is pale rufous in color similar to mexicana.
This was noted by Stone (1908), who stated that mehleri
“is indistinguishable from mexicana above, and differs below only in the
greater amount of black shading on the rectrices; the greatest difference is
found in the much larger bill”. Given that the very rufous coloration of mexicana
is one of the primary characters suggesting species status for this taxon, this
is of particular interest. Although proposal 2022-B-11 highlighted the similar
pale rufous plumage of mexicana and pallescens of eastern Brazil,
no specimen photos of mehleri were included in
that proposal. The similar pale rufous coloration of mehleri
and mexicana is readily apparent in photos, although the undertail of mehleri is darker overall, being more similar to
other taxa in the cayana group in this regard. Photos of mehleri from northern Colombia:
https://macaulaylibrary.org/asset/206165711
https://macaulaylibrary.org/asset/366888881
https://macaulaylibrary.org/asset/393722091
Another
critical issue in overlooked in 2022-B-11 are differences in orbital skin
color, something noted by Pam Rasmussen in her WGAC proposal and described in
detail by Fitzgerald et al. (2020), but of course not apparent in specimens. In
fact, this character might be a much better indicator of species limits in the
group than overall plumage coloration, the latter of which seems to vary
considerably based on climate. Based on Schulenberg et al. (2007), Restall et
al. (2007), Fitzgerald et al. (2020), and available photos online, variation in
orbital skin color is as follows: blue-gray in mexicana; greenish-yellow
in thermophila, nigricrissa, mehleri, circe, and insulana;
and red in mesura, cayana (of the Guiana
Shield), and all remaining South American taxa. Based on photos, it appears
that populations with red orbital skin (mesura
and cayana) approach those with greenish-yellow orbital skin (nigricrissa,
circe, and mehleri)
in multiple places with very abrupt turnover. These areas mostly correspond
quite closely to the subspecies turnovers noted by Chapman (1917). These
include in the southern Magdalena Valley near Neiva (greenish yellow mehleri to the north, red mesura
to the south/east), the Rio Orinoco in Venezuela (greenish yellow circe on the left bank, and red cayana on the
right bank), and perhaps somewhere across the Rio Meta in the dry llanos Orientales of Colombia. The two (here mehleri
and mesura) also appear to turn over within a
few kilometers along the eastern flank of the eastern Andes near Yopal, Casanare, Colombia: https://macaulaylibrary.org/asset/285186601 versus https://macaulaylibrary.org/asset/586198261. eBird photos from
Casanare department, Colombia in the dry llanos show a mix of red and
greenish-yellow orbital rings in a patchwork, raising the possibility of local
sympatry. We have found just one individual (from adjacent northern Meta
department) that appears to show some green in an otherwise red orbital ring,
which would argue for some limited hybridization in this area: https://macaulaylibrary.org/asset/217105071https://macaulaylibrary.org/asset/217105071. There is also abrupt turnover in this character within a
few kilometers across low Andean passes near Loja in southern Ecuador (here nigricrissa
and mesura). See https://macaulaylibrary.org/asset/518051361 versus https://macaulaylibrary.org/asset/600766311.
This
would all suggest that if a split is implemented, the division of only thermophila
and nigricrissa from cayana is not a good course of action. In
fact, we suggest based on orbital ring color and what appear to be very sharp
turnovers between populations with red vs greenish-yellow orbital rings, that a
group comprised of circe, mehleri, insulana, thermophila
and nigricrissa could be split from P. cayana. In this case, the
northern species would be either P. circe or P.
mehleri, both described by Bonaparte in 1850,
rather than P. thermophila Sclater, 1859. Because Bonaparte (1850)
described circe and mehleri
in the same publication, a first reviser action would likely be required to
establish priority; we will refer to this species as P. “circe”
through the rest of this proposal. Bonaparte’s description of these
taxa is here:

To
provide a better visualization of where these taxa come into contact in
Colombia, below is the eBird abundance map (https://science.ebird.org/en/status-and-trends/species/squcuc1/abundance-map), with a red line
approximately delineating the cayana and “circe”
groups based on the distributional statements above. The abundances do seem to
match the number of eBird records in these regions, so are likely a decent
representation of the distribution. However, it would be great to get some
insight on this issue from Colombian and Venezuelan ornithologists who are more
familiar with this species in these potential areas of contact. If implementing
this split, the range boundary between P. “circe”
and P. cayana would be approximately as such:
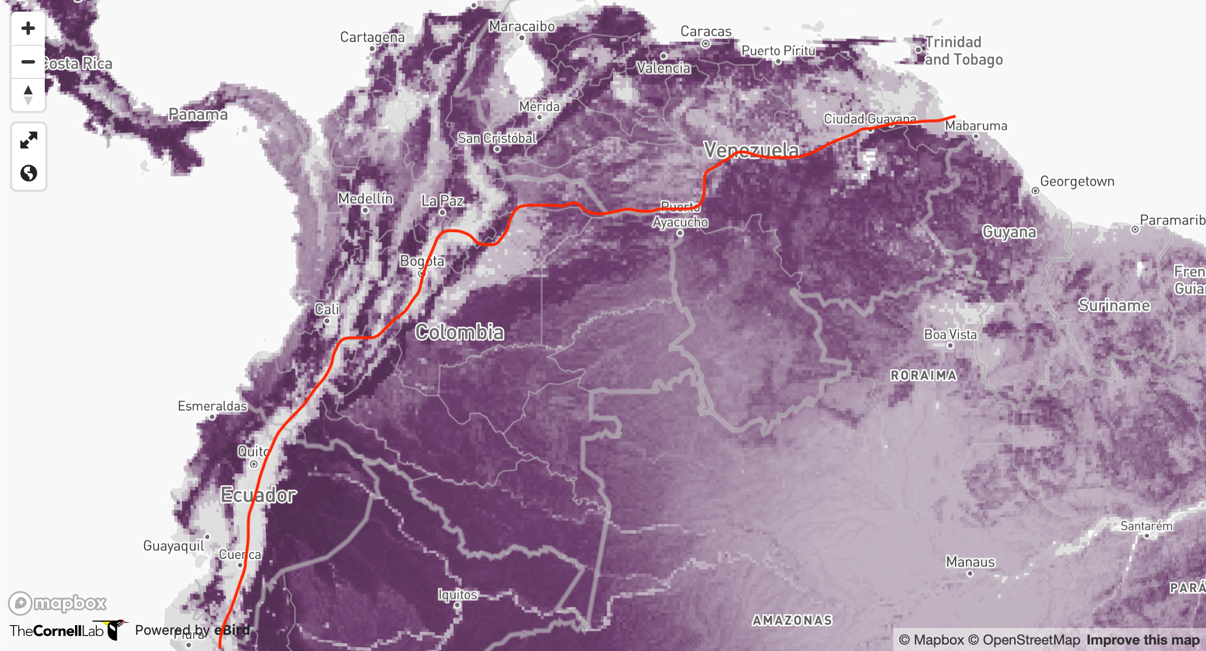
In
reading the older literature on this group there is a bewildering number of
synonymies for each taxon, which is confounded by multiple taxa described from
“Bogota” skins, and multiple examples of a name being applied to different
populations by different authors. Much of this was sorted out by Chapman (1917)
and Junge (1937) but we think some errors persist. As
an example of this confusion, Stone (1908) applied mehleri
Bonaparte, 1850, to the Central American populations (now considered thermophila)
based on Sclater’s (1860) determination that the type locality was in fact
“Central America”, not “Santa Fé de Bogota” as
originally given by Bonaparte. Chapman (1917) then applied columbiana
(type locality Cartagena, Colombia) to the northern Colombian population,
considering mehleri Allen, 1900 (type
locality Santa Marta, Colombia), as a synonym, apparently overlooking mehleri Bonaparte, 1850. Later authors (e.g.,
Fitzgerald et al. 2020) applied mehleri
Bonaparte, 1850, to the populations of coastal northern Colombia and Venezuela
(i.e., columbiana of Chapman 1917). We mention this because we have not
undertaken a thorough review of all synonymies for these taxa, and trust that
later authors (e.g., Fitzgerald et al. 2020) have resolved these issues
satisfactorily, such that if these taxa are split the correct names are applied
to the daughter species.
One
issue that we have attempted to clarify involves the type localities of circe and mehleri.
Junge (1937) sorted out these type localities by
reviewing the collecting localities on the tags of the type specimens. In
contrast to earlier authors (see previous paragraph), he reported that the type
of circe was collected in Caracas,
Venezuela, and mehleri in Cartagena, Colombia.
Both of these localities contain pale rufous birds with greenish-yellow orbital
rings, so can be confidently associated with the northern group, not with the cayana
group, based on orbital ring color. Phelps and Phelps (1958) thought that
the type locality of circe was likely
Mérida, Venezuela, and reported the distribution as being south of Lago
Maracaibo, which seems to be the basis of the distributional statement in
Fitzgerald et al. (2020). However, Junge (1937)
compared the type of circe (from Caracas) to
specimens collected “south of Lago Maracaibo” and concluded that they were
similar enough to be considered same taxon. So, we suspect that it is circe that is found from western Venezuela (near
Lago Maracaibo) as far east as the Delta Amacuro. Subspecies mehleri would then be restricted to northern Colombia
and the Magdalena Valley.
Effect on SACC area:
Splitting
“circe” from cayana would result in one
additional species for the SACC area. Splitting mexicana from cayana
would not result in any additional species for the SACC area, as mexicana
is extralimital. However, we think that it is still worthwhile for SACC to
consider this split, as it would be better to consider species limits in the
complex as a whole, based on current information.
Please
vote on the following issues:
A.
Treat Piaya mexicana as a separate species from P. cayana
B.
Treat Piaya “circe” (including thermophila,
nigricrissa, mehleri, and insulana)
as a separate species from P. cayana
Recommendation:
This
is clearly a borderline case with suboptimal data [and a potential
nomenclatural issue], but we tentatively recommend a YES on both A and
B.
The
split of mexicana would be based on the mitochondrial genetic
differences, consistent plumage differences, morphometric differences, possible
sharper call notes, and narrow contact zone with thermophila. This is
the treatment recommended by Sánchez‐González et al. (2023).
This contact zone does appear to be narrow and occurs across a sharp ecotone,
albeit with a few intermediates. However, there are still no formal analyses of
vocal or plumage data. The vocal information mentioned above does seem to indicate
a sharper, higher-pitched call note in mexicana, but it is unknown
whether these differences are diagnosable, if they’re affected by the level of
agitation of the bird, or if they’re relevant in playback trials. The plumage
differences between mexicana and
thermophila are readily apparent visually (especially the rufous undertail
of mexicana), but the overall pale rufous plumage coloration is repeated in
other taxa such as mehleri and pallescens.
The morphometric data from Sánchez‐González et al. (2023)
for mexicana vs. thermophila do show average differences between
the two groups, but with overlap. The data show that the longer tail of mexicana is closer to being diagnostic
versus thermophila than are other characters (i.e., less
overlap in the box plots). However, splitting mexicana from cayana, and not splitting thermophila, would render cayana paraphyletic for
the mitochondrial gene tree. This may not be an issue, given gene tree /
species tree issues, but nuclear DNA data would be preferable. There are also
little data on gene flow across the contact zone. If the Nm
values in Sánchez‐González et al. (2023) are reliable (which we
posit that they are not), there is little gene flow across the contact zone (Nm = 0.05). In short, there appear to be diagnosable differences across a
small contact zone, but it is not clear whether these differences correspond to
biological species.
In
contrast, as noted above, there are still many unanswered questions regarding
the contact zone between the “circe” and
cayana groups. Most importantly, there are no genetic samples from
potential contact zones in Venezuela, Colombia, or Ecuador. However, given the
available data, splitting “circe” and
mexicana would maintain monophyly for the mitochondrial gene tree and split
two clades that are 1.2 – 1.7 million years divergent. The data from Smith et
al. (2014) suggest that there is a mitochondrial clade in northern Peru that is
distinct from the rest of South America, and whether this clade is the same as
the one found in Colombia is unknown. We think that the critical data are from
the very abrupt turnover between taxa with red orbital rings from those with
greenish-yellow orbital rings. Analyses of these contact zones do seem critical
to determining species limits, but we think the data at hand tip the scales
towards valid species. However, the sonograms of the thermophila and
cayana groups shown above look fairly similar to us. There is also
still the issue of the extensive plumage variation within the cayana group
even if the “circe” group is split, with pale
bellied and pale rufous taxa (e.g., pallescens) and dark-vented (e.g., macroura)
taxa that at least superficially resemble the Middle American and northern
South American taxa.
The
nomenclatural issue of circe and mehleri is problematic. As stated above, because circe and mehleri
were published simultaneously, a first reviser action would be necessary to
decide which species name would apply if this northern group were split from P.
cayana. We think it is worth considering this novel species treatment given
the available data, but it may be worth waiting until for a publication to sort
out the nomenclatural issues before implementing the split.
The
way that the voting options are structured, there are a few other possible
voting solutions. The first is a YES on A and a NO on B, which would render P.
cayana paraphyletic in the mitochondrial gene trees. The other option would
be a NO on A and a YES on B, which would prioritize the deeper split in the
mitochondrial gene tree and the more obvious difference in orbital ring color
(the blue-gray orbital ring of mexicana is somewhat more similar to the
greenish-yellow of thermophila). Note that if this latter option is
adopted that the species would be P. mexicana (Swainson, 1827), which
has priority over circe Bonaparte,
1850.
Note
also that the WGAC is considering splitting just thermophila and nigricrissa
from the remainder of the South American taxa. We do not consider this a viable
solution to this taxonomic problem, and are not including it as a voting
option.
If
any of these splits are adopted,
an English name proposal should be drafted to address the new names, preferably
in coordination with the NACC. We tentatively recommend Mexican Squirrel-Cuckoo
for mexicana, following Chapman (1917). Although a bit of a mouthful,
Chapman (1917) used Central American Squirrel-Cuckoo for thermophila (although
nigricrissa occurs in South America), but this name would not be
appropriate for the more widespread P. circe.
One option, though not ideal, could be Northern Squirrel-Cuckoo for “circe” and Southern Squirrel-Cuckoo for cayana.
South American Squirrel-Cuckoo could also work for cayana.
Literature Cited:
Binford, L. C. 1989. A
Distributional Survey of the Birds of the Mexican State of Oaxaca.
Ornithological Monographs 43. American Ornithologists’ Union. Washington, DC,
USA.
Bonaparte, C.L. 1850.
Conspectus Generum Avium. Lugduni
Batavorum. Apud E. J. Brill, Acadamiae
Typographum.
https://www.biodiversitylibrary.org/item/141467#page/186/mode/1up
Chapman, F.M. 1917. The
distribution of bird-life in Colombia; a contribution to a biological survey of
South America. Bulletin of the American Museum of Natural History. Volume 36. https://www.biodiversitylibrary.org/item/117614#page/407/mode/1up
Cory, C. B. 1919.
Catalogue of birds of the Americas, part II. Field Museum of Natural History
Zoological Series Vol. XIII. Chicago, USA.
Fitzgerald, J., T. S.
Schulenberg, and G. F. Seeholzer. 2020. Squirrel Cuckoo (Piaya cayana), version 1.0. In Birds of the World (T. S.
Schulenberg, Editor). Cornell Lab of Ornithology, Ithaca, NY, USA. https://doi.org/10.2173/bow.squcuc1.01
Junge, G.C.A. 1937. On
Bonaparte’s types of the cuckoos belonging to the genus Piaya. Zoologische Mededelingen
19:186–193.
Navarro-Sigüenza, A.G.
and A.T. Peterson. 2004. An alternative species taxonomy of the birds of
Mexico. Biota Neotropica 4(2):1–32.
http://www.biotaneotropica.org.br/v4n2/pt/fullpaper?bn03504022004+en
Peters, J. L. 1940.
Check-list of birds of the world. Vol. 4. Museum of Comparative Zoology at
Harvard College.
Phelps, W.H., and W.H.
Phelps, Jr. 1958. Lista de las Aves de Venezuela con su
Distribución. Tomo II, Parte I. No Passeriformes. Editorial Sucre, Caracas.
Restall, R., C. Rodner,
and M. Lentino. 2007. Birds of northern South America; an identification guide.
Volume 1: species accounts. Yale University Press.
Ridgely, R.S., and P.J.
Greenfield. 2001. The birds of Ecuador. Vol. 2. Cornell University Press.
Ridgway, R. 1916. The
birds of North and Middle America. Part VII. Bulletin of the United States
National Museum. No. 50.
Sánchez‐González,
L.A., H. Cayetano, D.A. Prieto‐Torres, O.R. Rojas‐Soto,
and A.G. Navarro‐Sigüenza. 2023. The role of ecological and
geographical drivers of lineage diversification in the Squirrel Cuckoo Piaya
cayana in Mexico: a mitochondrial DNA perspective. Journal of Ornithology
164:37–53. https://doi.org/10.1007/s10336-022-02008-w
Schulenberg, T.S., D.F.
Stotz, D.F. Lane, J.P. O’Neill, and T.A. Parker III. 2007. Birds of Peru,
revised and updated edition. Princeton University Press.
Stone, W. 1908. A
review of the genus Piaya Lesson. Proceedings of the Academy of
Philadelphia 60(3):492–501. https://www.jstor.org/stable/4063306
Swainson, W. 1827. A
synopsis of the birds discovered in Mexico by W. Bullock, F. L. S., and H. S.,
Mr. William Bullock. Philos. Mag. (New Series) 1: 364–369, 433–442. https://www.biodiversitylibrary.org/item/53108#page/456/mode/1up
Whitlock, M., and D.
McCauley. 1999. Indirect measures of gene flow and migration: FST≠1/(4Nm+1).
Heredity 82:117–125. https://doi.org/10.1038/sj.hdy.6884960
Oscar Johnson,
September 2023
Comments from Remsen: “NO. Highly conflicted on this
one. Borderline indeed. If the data assembled in the proposal were in
the form of a peer-reviewed publication, I would vote YES because I think it
more closely approximates species limits than does the current broadly defined,
Peters-based single species.
“A thorough study
that sampled all taxa might reveal some conflicts with a three-species
taxonomy, and it is always easy to require more data before making splits in a
complex like this one. But a “do no harm” conservative approach might be the
wisest thing to do in a case like this given the data gaps. We already know that basing taxonomy on mtDNA
gene trees is unwise, so I don’t trust the basics of the tree typology. And the contact zones outside of the mexicana-thermophila
one are poorly characterized. And then
there’s the circe-mehleri nomenclature
problem.
“Given the
absence of any sign of free gene flow between extralimital mexicana and thermophila,
I personally would consider that sufficient evidence for a species split, but
that is out of the SACC realm. Given the
orbital ring color differences outlined in the proposal, I think it also might
be safe to extrapolate from the mexicana-thermophila situation to
the South American situation. If all
this were summarized in a publication rather than NACC + WGAC proposals, I
would favor a YES vote, especially because that would also provide the first
revisor opportunity.”
Comments from Robbins: “NO. This
is indeed a difficult situation to evaluate, at any level. As Oscar summarizes
in paragraph two of the recommendation, a number of issues need to be clarified
with just splitting off mexicana.
I’m not convinced that plumage differences have any relevance in species
limits, and I even wonder if orbital ring color is important. Moreover, as Oscar points out the vocal
comparisons may not have been standardized, i.e., comparing analogous calls
under similar conditions, and I agree they not only look similar in a
spectrogram but sound similar. It should
also be noted that only one of several vocalizations that Piaya give
have been looked at. Finally, I would like to see a nuclear DNA data set that
helps guide the way forward. Thus, for now, I vote NO.”
Comments
from Lane:
“NO to both A and B. Having some personal experience
with both Middle American/trans-Andean groups, and extensive experience with cis-Andean
groups, the main characters that define them are not strong enough for me to
see them as species-level. The cis/trans split seems like it is the one we
should be looking at first, as it clearly is the older split, and the orbital
color is a character that doesn't seem as affected by Gloger's Rule as the
plumage, but even there, I'm underwhelmed by the vocal differences, which to me
suggest that these groups simply aren't that divergent. I wonder if work in the
Llanos of Colombia and Venezuela might elucidate how the two groups interact?
As indicated in the proposal, there has been effectively no real effort yet to
quantify vocal differences, and whereas the mitochondrial groups certainly
indicate clades that could be worthy of recognition, my mind doesn't register
them as so distinctive as to warrant species status.”
Comments from Zimmer:
“A. Treat
Piaya Mexicana as a separate species from P. cayana. NO.
“B. Treat Piaya “circe” (including thermophila, nigricrissa, mehleri, and insulana)
as a separate species from P. cayana. NO.
“This
is a tough call for me, because, from the evidence presented in the Proposal, I
suspect that the proposed three-way split is actually the most reflective of
the relationships of these various taxa.
The abrupt turn-over in orbital ring color between the cis-trans groups
is particularly compelling to me, as is the absence of any sign of free gene
flow between mexicana and parapatric thermophila. However, I
agree with Mark that the plumage differences and tail length differences,
although pronounced between some taxon-pairs, when considered as a whole, are
all over the map, and may not have any relevance in species limits. After all, plumage distinctions of tail
pattern, belly/vent coloration, and overall color saturation of upperparts and
underparts, and tail length of P. melanogaster seem to overlap or at
least closely approach, one or more named subspecies of “Squirrel Cuckoo”, but
bare parts colors and head/facial pattern are dramatically different (which, in
my mind, lends further significance to the cis-trans orbital skin color
differences in the cayana-group).
The larger issue in my mind, is the lack of any well-sampled vocal
analysis that compares clearly homologous vocalizations. As Mark notes, members of this complex have
multiple types of vocalizations, and, although, as alluded to by both Mark and
Dan, the vocal distinctions may not be impressively obvious on first
reflection, I suspect that a comprehensive analysis (both quantitative and
qualitative) will reveal some distinctions that are concordant with the
proposed splits. Looking
again at P. melanogaster, there are several different types of
vocalizations in that species as well, and most/all are similar enough to be at
least recognizably analogous to the various vocalizations within cayana
(sensu lato), and yet the birds themselves, as well as a rigorous vocal
analysis, would have little trouble making the distinction. I’ve also noticed a seeming difference in the
frequency of usage of homologous vocalizations between different populations of
Squirrel Cuckoos, particularly with respect to Atlantic Forest populations
versus Amazonian and Central American populations, which further underscores
the need for a vocal analysis that considers the entire range of the complex. Taken in sum, that leaves me as a reluctant
NO vote for now, but looking forward to a published analysis of vocalizations
that, I predict, will flip my vote to a YES.”
Comments
from Bonaccorso:
“A.
NO. But very close to a YES. The mitochondrial tree
from Sanchez-Gonzalez et al (2023), based on a good sampling of P.c.
thermophila and P. mexicana (at least for Mexico) and is very well supported, but I
think nuclear evidence should provide the definitive answer. Although
morphological measurements overlap, plumage variation seems clear, especially
regarding tail color. I agree that voice comparison needs to be standardized.”
“B. NO. It is too risky to make any decision without proper
genetic sampling of all the populations, and enough samples from each
population. For these, there are no formal morphometric or vocal analyses and
no comprehension of the potential contact zone.”
Comments from Claramunt: “NO. I’m persuaded by de evidence
for species status for mexicana. Here we have two populations with clear
phenotypic differences in plumage color, tail length, and skin color, that are
reciprocally monophyletic in their mtDNA genealogies, and show an extensive
contact zone with barely any evidence of potential hybridization. This pattern
would be highly implausible if the two taxa were not largely reproductively
isolated. However, the genetic evidence suggests that the separation of mexicana
would require further splits in the other taxon in order to avoid a paraphyletic
group. But the situation in this other group is complex. I agree with Van in
that situations of this complexity require a peer-review publication. I would
need to see maps with the distribution of subspecies and, more importantly,
maps of distributions of characters. And the nomenclatorial issues would need
to be worked out. I think there is great potential for this to become a
three-way split with periorbital skin color separating two South American
forms. But I would like to see the evidence more clearly.”
Comments
from Stiles:
“NO for now – this species could be split into two or more species, but the
bewildering number of noncongruent characters and holes or uncertainty in the
distributional limits of some forms and hence, in locating possible areas of
contact all add up to a puzzle with too many missing pieces. Just how one
potentially useful feature, eyering color, needs to be looked at with more
detail (hopefully, I may be able to look at the distributions of a number of
Colombian specimens here) – but for now, no clear decision seems possible.”