Proposal (999) to South
American Classification Committee
Treat Sakesphorus pulchellus as
a separate species from S. canadensis
Note:: This is one of
several situations that the IOU Working Group on Avian Checklists has asked us
to review because published treatments of the species differ, and WGAC has to
pick one. BirdLife International (del
Hoyo & Collar 2014) now treat these two as separate species. This is a complex situation about which I
have no first-hand knowledge, so my goal here is to lay out as many facts as
possible that might be of potential relevance.
Background: As
currently defined, Sakesphorus canadensis consist of 6 subspecies,
although Zimmer & Isler (2003) noted that the validity of some was
questionable due to apparent clinal variation:
• S. c. pulchellus (Cabanis & Heine, 1860) – n.
Colombia and nw. Venezuela
• S. c. intermedius (Cherrie, 1916) – e.
Colombia, s. Venezuela, n. Brazil
• S. c. trinitatis (Ridgway, 1891) – ne.
Venezuela, Trinidad, Guyana
• S. c. canadensis (Linnaeus, 1766) – Suriname, n. French
Guiana
• S. c. fumosus Zimmer, 1933 – s.
Venezuela in s. Amazonas
• S. c. loretoyacuensis (Bartlett 1882) – riverine
distribution in extreme se. Colombia, ne. Peru, and w. Amazonian Brazil
Restall et
al. (2006), however, also recognized two additional subspecies: paraguanae Gilliard, 1940, from coastal NW Venezuela
(type locality from Paraguaná Peninsula) and phainoleucus Todd, 1916, from ne. Colombia and nw.
Venezuela (type locality Rio Hacha). Restall et al. gave characters for both of
those in addition to characters for intermedius and trinitatis. This must be taken seriously because Restall
et al. based their conclusions on the world’s best collection of Venezuelan
birds, the Phelps Collection. Cory and
Hellmayr (1924) treated phainoleucus as a
synonym of pulchellus with the following footnote from Hellmayr, which
implies that Todd actually then agreed with him:
“Birds from the Goajira Peninsula and nw. Venezuela (Rio Aurare, se. of Altagracia, Zulia; Barquisimeto, s. Lara)
have slightly larger bills, more white on forehead, the sides of the head
mainly white, and less black on the under parts, this
color being, on
the throat, sometimes nearly concealed by the white apical portions of the
feathers. As, however, about fifty percent of the specimens are
indistinguishable from pulchellus, of nw. Colombia, I agree with E. W.
C. Todd that the recognition of phainoleucus
is of no practical advantage.”
Peters
(1951) listed paraguanae as a synonym of phainoleucus, and del Hoyo & Collar’s
supplemental material (see below) treated them both as synonyms of pulchellus
because they “appear to intergrade with pulchellus.” As stated, that makes no sense to me –
parapatric subspecies should intergrade at the contact zone. If they didn’t, it would be prima facie
evidence for a hard barrier to gene flow and hence full species rank.
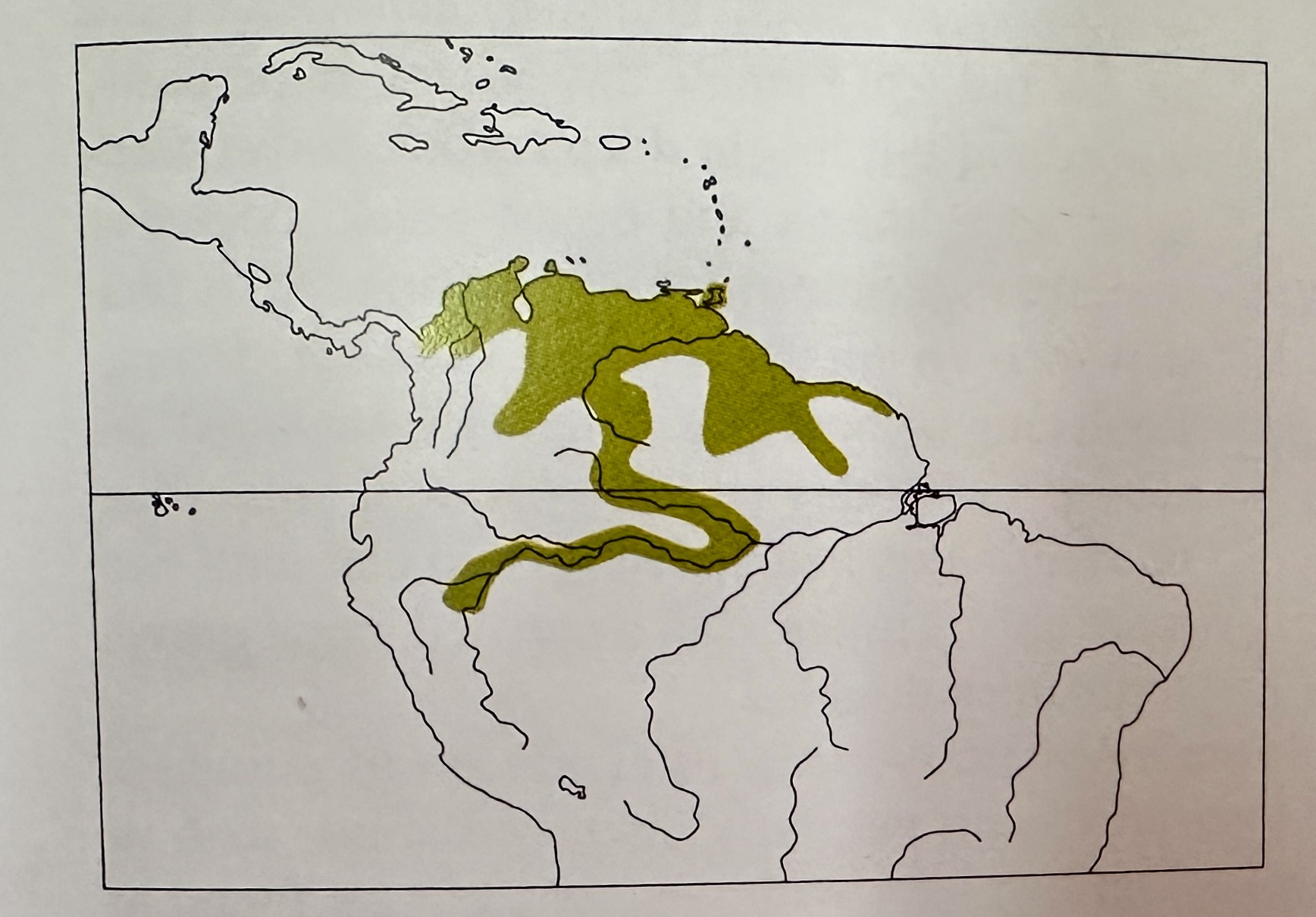
Here is the composite distribution of S. canadensis from
Zimmer & Isler (2003):
Here
is the terrific plate by Hilary Burn from Zimmer & Isler (2003), which
illustrates canadensis, pulchellus, loretoyacuensis, and fumosus. Trinitatis and intermedius (not
shown) are “very similar to nominate canadensis” (Zimmer & Isler
2003), and Cory & Hellmayr (1924) basically provided sufficient evidence in
their long footnote to treat them as clinal variation within a more broadly
defined canadensis; in retrospect, I (in Dickinson & Christidis 2014)
perhaps should have treated them as synonyms of the nominate subspecies because
Cory & Hellmayr’s (1924) qualitative assessment of them was sufficient to
put the burden of proof on maintaining them as separate taxa, and they treated intermedius
as a synonym of trinitatis. Note
that Zimmer & Isler (2003) didn’t even try to describe the differences
between intermedius and trinitatis versus the others because,
evidently, they were so subtle, although see Restall et al.’s subsequent
diagnoses of these subspecies. Peters
(1951) retained intermedius, perhaps on the basis of an analysis I
haven’t found. Cherrie (1916) described intermedius
as follows (and note that the paleness would be “in the direction” of pulchellus):
“Birds from the middle Orinoco, from Ciudad Bolivar,
and beyond, are intermediate in general color between the Cayenne birds and
those from Trinidad, the Orinoco delta, and Caura River points, being as a
series, at once distinguishable by their paler coloring both above and below.
This pale coloring is perhaps more marked in the females, but is very evident
in the males also when compared as a series. So characteristic does this paler
form seem of the middle Orinoco region that I would designate it as … HYPOLOPHUS CANADENSIS INTERMEDIUS subsp. nov.”
Based
on this plate, it looks to me that we have taxa in which both male and female
plumage roughly follow Gloger’s Rule, with the taxon from the driest area (pulchellus)
the palest, and the one from perhaps the wettest area (fumosus) the
darkest.

Here
is our current SACC note:
2h. The northwestern subspecies pulchellus
differs vocally from the others and may merit recognition as a separate species
(Zimmer & Isler 2003).
As far as I can tell, Ridgway
(1911) was the first one to treat pulchellus as a subspecies of S.
canadensis, and no rationale was provided nor does his synonymy indicate a
previous publication that had done that.
I can see the rationale, however, in the plate above – without checking
specimens, pulchellus superficially looks like a canadensis that
shows white streaking and freckling in the face, crown, and throat – other than
that, those two appear to resemble each other more than either does to loretoyacuensis
or fumosus. Cory & Hellmayr
(1918), Peters (1951), and all subsequent classification except del Hoyo &
Collar (2016) have treated them as conspecific.
Zimmer (1933) described fumosus
from 6 specimens taken near of Mt. Duida in Venezuela; he noted that two of the
males from “Río Cassiquare” [Casiquiare] not as
solidly black as the type in that “the irregular whitish marks on the sides of
the breast and belly slightly larger.”
He also looked extensively at variation in canadensis, and his
comments are worth noting here in full because (1) they illustrate in detail
the complexity of the situation, and (2) they underscore the problem of
assigning Tobias et al. plumage scores to taxa.
I suggest at least a skim, although it doesn’t get much better than a
full dose of Zimmer’s style:
“I am not able to adopt unreservedly the
arrangement proposed by Hellmayr (Field Mus. Nat. Hist. Publ., Zool. Ser.,
XIII, pt. 3, p. 53, 1924) which assigns all of the Venezuelan birds (except
pulchellus of the Lake Maracaibo region) to trinitatis; yet, with fewer
skins than the series examined by Hellmayr, I would hesitate to make a counter proposal
were it not for certain new material at hand which helps to explain some of the
puzzling factors in Hellmayr's arrangement.
“With the establishment of typical canadensis
in French and Dutch Guiana, the birds from Trinidad are recognizable under the
name trinitatis. I have only two males from British Guiana, one of
which, labeled "Demerara," is not unlike Trinidad males while the
other, collected by Alexander and possibly from the eastern portion of the
country, is much like true canadensis. The difference between the males
is small and, without females from various parts of British Guiana, it is
impossible to say whether or not both forms occur in this country.
“In the Orinoco Delta region and in the
former state of Bermuidez (now Anzoátegui and
Monagas), the birds are very like the Trinidad examples.
“Farther up the Orinoco, at Ciudad
Bolivar, Caicara, and the Rio San Feliz, there is a
prevailing tendency toward lighter coloration than is shown in the delta
region. The males are brownish on the back, rather than grayish, but the tone
is light, and the lores are decidedly whitish. The under tail-coverts are
largely white, sometimes grayish subterminally but without a strongly blackish
area in that position such as occurs in canadensis and trinitatis.
The sides and flanks are light gray or even whitish, in reduced contrast to the
white area bordering the median black stripe. The females also are pale brown
on the back, and are light rufous on the crown, pale ochraceous below, with
only moderately heavy streaking on the breast, and with the belly distinctly (though
restrictedly) white in the middle. The same style of coloration, possibly a
trifle warmer, is exhibited by birds from the Rio Surumú,
Brazil, an affluent of the Rio Cotinga. The region of the Surumui
is largely savanna country, I am informed by Messrs. Tate and Carter, who
visited the locality, and savanna occurs at places on the top of the Pacaraima
Range and at the headwaters of the Rio Caroni in Venezuela and it may extend,
at least brokenly, down to the middle stretches of the Orinoco. Consequently it
seems entirely possible that a light-colored race may exist in these savannas.
Since the Caicara bird has been named intermedius
by Cherrie, that name would be available for such a pale subspecies, if it can
be satisfactorily maintained.
“Birds from the state of Falcon are
neither typical trinitatis nor the Caicara
form but probably are nearer the latter though they are slightly darker. The
under tail-coverts are without blackish subterminal areas and the lores of the
males are rather extensively whitish. Possibly these birds should be considered
as intermediate between trinitatis and pulchellus which latter
form inhabits the nearby state of Lara, but, even if so, the similarity to intermedius
may necessitate their reference to that subspecies. An additional character
noted in the two males from the state of Falc6n, but not observed in skins from
other regions nor in Falcon females, is a small whitish area on the inner webs
of the tail feathers at their extreme base. Its significance is not clear. In
any case, material must be examined from the region between Caicara
and the state of Falcon to determine the possible continuity of range. Since
the region is one of savannas, direct connection is not unlikely. Nevertheless,
Hellmayr and Seilern (Arch. Naturg., LXXVIII, A (5),
p. 119, 1912) found three males from San Esteban, Carabobo, to be more grayish,
less rufous, above than others from British Guiana, Trinidad, and the Rio
Branco, Brazil, being like skins from the Caura region and San Fernando de
Apure!
On the Río Caura, a different type of
coloration is encountered which is not that of Caicara
and Ciudad Bolivar although the Caura empties into the Orinoco between these
two places. Judging by the darker hues, the Caura birds are inhabitants more of
forests than savannas and, from available accounts, the Caura is marked by this
type of habitat. Some relationship to the forest-inhabiting fumosus is, therefore,
to be expected. Four males from as many localities (Río Mato, Suapure, Maripa, and La Uni6n)
all have the white stripes bordering the median black area of the under parts
virtually obsolete, being dull and grayish and not distinguishable as
sharply-defined white; the lower belly is quite sooty, not white. The
metacarpal border of the under wing-coverts is broadly black in the Río Mato
male, with white tips in the Maripa skin, intermediate
in the other two; the under tail-coverts have blackish bases and relatively
narrow white tips; the male from La Uni6n has the lores quite black, though the
three other males have much white in this region. All these tendencies are in
the direction of fumosus. The back is rather plain, without the heavy
streaks of fumosus but of a darker tone than in intermedius. An
additional character of doubtful significance is the decided reduction of the
white spot on the lateral margins of the outer pair of rectrices. Instead of
the customary broad patch reaching from the shaft to the margin, there is only
a narrow marginal streak, rarely supplemented by a small oval spot in the
middle of the web. A young male has the patch of exceptionally large size,
connecting on the left rectrix with the white at the tip of the feather. In the
Caura females the patch is of the regular size or but slightly reduced, and, in
addition, the general color of both upper and under parts is as near to that of
female intermedius as to that of fumosus, being intermediate
between the two, as in the males. The general impression left by the Caura
birds of both sexes is that of intermediates between intermedius and fumosus,
not definitely referable to either.
“In the neighborhood of the upper Orinoco,
above Caicara, from Maipures
to the falls of the Atures at Ayacucho, another
definite change of color and pattern is found which bears little relation to
the Caura series. The males from this region are even darker and more rufous
brown on the back than the Caura males with the added features of rather prominent
dusky streaks and an evident, though very small, concealed patch of white on
the mantle. The white patch on the outer margins of the outer rectrices is not
reduced in size but the lores are noticeably whitish. However, the under parts
have not lost any of the broad white areas but rather have this white more
decidedly in evidence than usual and the lower belly is white, showing no
approach toward fumosus in these respects. I have no females from this
part of the Orinoco, but a young male from the "Upper Orinoco"
(judging by the collector's dates, not far from Maipures)
is very like the young male from the Caura (Maripa)
and, like it, has unusually extensive white on the outer margins of the outer
rectrices, but is a little duller on the mantle. [Curiously enough, a young
male of intermedius from the Rio Surumú,
Brazil, and one from Ciudad Bolivar, Venezuela, also have the white spots of
tip and outer margin of the outer rectrices continuous, with a small subterminal
spot of dusky on the outer web, and a young female of trinitatis from
Las Barrancas, Rio Orinoco, has the same continuity without any dusky spot, as
in pulchellus. An occasional skin of pulchellus shows a
subterminal dusky spot and one male from La Cienega, Santa Marta, has the
marginal patch connected with the terminal spot only by a very narrow line on
the outer margin of the outer web.] There is a tendency toward the development
of white not only on the lores, as mentioned, but also on the forehead and
superciliary region (as well as in the malar region where it appears frequently
in other forms). The under tail-coverts are grayish as often as sooty and are
quite broadly tipped with white which conceals the darker basal portions. The
nearest affinity, in several of’ these respects, is pulchellus which
occurs southeast of Lake Maracaibo though on the far side of the cordillera
which separates the drainage of this lake from the rivers flowing to the upper
Orinoco.
“A single male from San Fernando de Atabapo, between Ayacucho and Mt. Duida, is more like fumosus
than are the Ayacucho birds, but it still has the white on the sides of breast
and belly and the streaked upper parts with more of brown than of black though
the brown is grayer and Jess rufous than in Ayacucho males. Resemblance is
apparent to some loretoyacuensis, which probably is due not to racial
consanguinity but rather to a parallelism reached in the transition from fumosus
to pulchellus or intermedius.
“The only other skin which needs special
mention is a male from Caracarahy, on the middle
stretches of the Rio Branco, Brazil. This bird is plain brown on the mantle, of
a darker hue than that of intermedius from the affluents of the upper
Branco; in other respects it resembles loretoyacuensis. Since it comes from a
locality in the region where the ranges of these birds must meet, it may be
considered as intermediate between them.
“In spite of the apparent regularity of
the variations on the Caura and at Ayacucho and Maipures,
and the impossibility of referring the respective series to one form or
another, I hesitate to name new forms from these two regions. Obviously fumosus,
as an inhabitant of forested areas, finds its way across the Pacaraima Mts. to
the upper Caura which is forested, and extends down that stream in somewhat
modified form, affected, probably, by some contact with the paler intermedius
of the savannas which is more prevalent to the east and northward.
“On the other hand, descending the
Orinoco, an earlier contact occurs with savanna-covered regions, and a
different modifying factor may exist in pulchellus some distance to the
northwestward, resulting, in any event, in a somewhat different combination of
characters as outlined above. Until more material is available from other
localities I can do no more than suggest the lines of possible relationship.”
New information:
Del Hoyo & Collar (2016)
treated pulchellus as a separate species (“Streak-fronted Antshrike”)
from canadensis and other 4 subspecies.
Unfortunately, their plate places the illustrations of pulchellus
and nominate canadensis on opposite sides of the page so that the similarities
in plumage are not emphasized. Del Hoyo
& Collar’s assessment using the Tobias et al. point scheme is as follows
(courtesy M. Iliff):
“Streak-fronted: https://birdsoftheworld.org/bow/historic/hbw/bkcant1/1.0/introduction:
Previously suggested as possibly deserving
of species status, but hitherto treated as race of S. canadensis.
Differs (in male) in its black hood shot with white (forecrown with white
streaks, white speckling on throat, head-side and supercilium) (2); upperparts
bright pale cinnamon vs dullish cinnamon-brown (or darker) (1); more white in
tail, with much larger white tips and all-white outer vane of outermost rectrix
(small white tips in other taxa) (2); more white on underparts (ns[1]); and
overall distinctive song, with slower pace and lower frequency at start, lower
maximum frequency (no overlap; 3), and last notes dropping vs rising in pitch
(3), with less acceleration (1). Proposed races phainoleucus
(Riohacha, in N Colombia) and paraguanae
(Paraguaná Peninsula, in NW Venezuela) appear to
intergrade with pulchellus, and are best rejected. Monotypic.”
As an aside, fumosus certainly would score more points based
on plumage vs. nominate canadensis than pulchellus does, and loretoyacuensis
also would likely to do so.
The vocal differences are based on Boesman (2016), who
documented substantial differences in the loudsong of pulchellus (N=6)
and “other races” (N=8): faster pace, last notes dropping in pitch, lower start
frequency, snarling notes at end, etc. I’m
a big fan of Boesman’s notes as catalysts to guide more thorough research, but
he pumped out 457 of these in a short time and just did not have the time to
get them into the shape that would be required for peer review. For example, It is not clear from Boesman
whether his 8 samples from “other races” included all 5 subspecies; he stated
that “song shows little variation among the different races, with the exception
of race pulchellus.” What would
be really critical is a comparison of songs from adjacent populations of trinitatis;
from range maps it would appear that there is a potential contact zone in north-central
Venezuela in the vicinity of Yaracuy-Carabobo and also south of Lake Maracaibo. In fact, here’s a footnote by Hellmayr from
Cory & Hellmayr (1924) that describes a specimen intermediate between pulchellus
and trinitatis:
“A single male from Catatumbo, sw.
of Lake Maracaibo, however, combines the general coloration above and the
extensive white tail-markings of pulchellus with the dark gray flanks
and the chiefly black sides of the head of trinitatis. More material is
required to prove the constancy of these characters or otherwise.”
However, I wonder if that is not in the range of intermedius
rather than trinitatis – the boundary between trinitatis and intermedius
is not clear to me.
Some sample recordings:
• pulchellus from n.
Colombia by Ross Gallardy: https://xeno-canto.org/353175
• trinitatis: from Aragua,
Venezuela, by Chris Parrish: https://xeno-canto.org/6209
• intermedius: from Apure,
Venezuela, by Joe Klaiber: https://xeno-canto.org/220949
• canadensis: from French
Guiana by Olivier Claessens: https://xeno-canto.org/42702
• loretoyacuensis: from Ilha
Anavilhanas west of Manaus by Thiago V. V. Costa: https://xeno-canto.org/14906; and from
Peru by Juan Díaz Alván: https://xeno-canto.org/87932.
• fumosus: from Roraima by
Jeremy Minns: https://xeno-canto.org/319232
My cursory cruise through the recordings supports everything that
Boesman said, especially the differences in the ending of the song. By the way, the differences in the song
endings of the pulchellus group were noted long ago by David Ascanio as
cited by Hilty (2003). On the other
hand, all the songs sound somewhat alike to someone like me who is not immersed
in antshrike song. Whether the
differences meet the Isler-Whitney criteria for species-level differences is
not obvious (to me).
New genetic data: My view on genetic data and
species limits is that unless they reveal non-monophyly, they are not taxonomically
definitive. Using comparative branch
lengths and genetic distance is problematic in my opinion. Harvey et al.’s (2020) suboscine
tree shows the following branching pattern, strongly supported, for Sakesphorus:

The salient points here for me are:
1. I
originally thought that that the Casanare sample referred to pulchellus
because comments from WGAC voters stated this.
However, I am grateful to Ottavio Janni (pers.
comm.) for pointing out to me that a sample from Casanare would represent intermedius,
of del Hoyo and Collar’s narrowly defined canadensis group, NOT pulchellus,
as indicated in private comments from WGAC members. Therefore, pulchellus sensu del
Hoyo-Collar is a paraphyletic taxon that also conflicts with the vocal data.
2. If one
puts stock in genetic distance, the depth of the node that marks the split of
the three Thamnophilus schistaceus samples is considerably deeper than
the one that the comparable node for S. canadensis sensu lato. Also, the node that marks the split of the five
T. murinus samples is at the same depth as the comparable node for S.
canadensis sensu lato. I only
included a portion of the tree just to include the first Thamnophilus
branch. Whether the canadensis
node better fits the current taxonomic ranks of species vs. subspecies in Thamnophilus
would require more work.
Discussion
and recommendation:
I’m going to try to get an antshrike expert to take my vote on this because I
can see this both ways and am conflicted.
If there were a study of the likely contact zone between pulchellus
and trinitatis in north-central Venezuela, then this would be a simple
problem – do they intergrade there or not?
The same applies to pulchellus vs. intermedius. Even a densely sampled set of recordings from
that area would show whether the song characters change abruptly or not. If there were an analysis of the vocal
differences using the Isler-Whitney criteria, then that would also provide an
objective answer. But as is, we have
none of that. All we really have is a
summary of an analysis of 14 recordings, possibly but not certainly of all the
subspecies, and a sum of plumage character differences of which most individual
characters are typical of subspecies-level, not species-level, differences. Further, pulchellus is much more
similar overall in terms of plumage to the adjacent subspecies of canadensis
(trinitatis and intermedius) than those two are to other
subspecies that del Hoyo & Collar included in canadensis sensu
stricto. The fatal flaw in the del
Hoyo-Collar treatment the genetic data (Harvey et al. 2020) conflict with the vocal
data similarities, and refute treating their treatment of canadensis as
a species. Finally, I feel reluctant to
make any decisions on this complex until a thorough, modern analysis of
geographic variation is presented. Badly
needed is a study of character variation.
Every author who has commented on plumage variation has mentioned some
degree of variability within samples from within even a single taxon. The actual boundaries between taxa are also
unclear, at least to me. This make me
uncomfortable. All of these issues constitute
sufficient reason, in my opinion, to proceed cautiously in any such decision,
and so I strongly recommend a NO vote on this one.
English
names: If the proposal passes, I think we should
have a separate proposal on English names. Streak-fronted is excellent. However, leaving the rest of canadensis
with the parental name Black-crested will lead to perpetual confusion, i.e. the
same name, Black-crested Antshrike, would apply to two different taxonomic
treatments. Broadly defined Sakesphorus
canadensis has been known as Black-crested Antshrike or Something
Black-crested Antshrike since the dawn of English names for South American birds
(e.g. Cory & Hellmayr 2024). The
range of pulchellus is large; although not as large as canadensis
sensu stricto, it is far from being a peripheral isolate. So, if we follow our guidelines (C.1), then we should
coin a new name, perhaps something like Black-fronted Antshrike to maintain as
much of the connections as possible.
References (see SACC Bibliography for standard
references)
Boesman, P.
2016. Notes on the vocalizations
of the Black-crested Antshrike (Sakesphorus canadensis). HBW Alive Ornithological Note 52: In Handbook
of the Birds of the World Alive. Lynx
Edicions.
Van Remsen, June 2024
Addendum
from Remsen:
This proposal suddenly became a lot easier thanks to Ottavio Janni, who pointed out to me that the genetic sample from
Casanare (llanos of Colombia) must refer to intermedius, of the del
Hoyo-Collar species canadensis, NOT to pulchellus. I had overlooked this because WGAC internal
discussion erred in assuming it represented pulchellus, and I didn't
catch that mistake. From Ottavio Janni: “I noticed that in
discussing the Harvey et al suboscine tree you say that "the two samples
of pulchellus cluster together", but the Colombian specimen is from
Casanare in the llanos, so shouldn't it be intermedius? This is what the
Hilty guide shows. The eBird photos from Casanare, a handful of which are mine,
also show birds that belong to the canadensis group on plumage (https://media.ebird.org/catalog?taxonCode=blcant4&mediaType=photo®ionCode=CO-CAS), although some birds have more of a grizzled instead of
uniformly black face (I guess that's why whoever described them named them intermedius?).
It would be good to clear this up, if indeed pulchellus and intermedius
cluster together I guess that would weaken the case for splitting pulchellus,
at least genetically, though maybe voice can make the case alone?”
_____________________________________________________________________________________________
Comments
from Stiles:
“NO for the reasons stated by Van: this whole complex of Sakesphorus appears
to be something like a maze of discordant characters of genetics, distributions
and color patterns that needs to be straightened out with a more detailed study
that definitely requires more data on vocalizations.”
Comments from Areta: “NO. I am torn
by this case, which seems to be solid, but for which more comparative
information is missing. The vocalizations differ noticeably, and Paul Schwartz
realized about that a long time ago (see here: https://macaulaylibrary.org/asset/61724).
“I think that pulchellus
will eventually be split from canadensis,
and it is a great candidate. As in other cases, if we were starting from
scratch, I would start by calling it a species. However, I would like to see a
full vocal study to help clear any doubt and to provide a more robust picture
of geographic variation in S.
canadensis as a whole. I stress that, biogeographically, numerous
species are restricted to arid N Colombia and N Venezuela. Everything makes
sense for a split here, but I want to see a more thorough assessment that goes
beyond the obvious. So, a
painful NO, until someone publishes a deeper study, that will
surely show that pulchellus
is a different species.”
Comments
from Robbins: “NO. Van has
underscored the issues with evaluating the complexity of these Sakesphorus
and that alone should raise concerns about making any changes. Moreover,
subsequently more information has been provided that has underscored issues
with the allocation of both genetic and vocal data to appropriate taxa (fide
Ottavio Janni, Gustavo Bravo, respectively). Thus, this has become a mess and clearly
needs to be sorted out before any taxonomic changes be made.”
Comments from Claramunt: “NO. I think Van’s
arguments are very valid. We have only spotty information on plumage and songs
(and DNA) from a complex that shows marked geographic variation (at least in
plumage). Climate and/or habitat seem to be playing a major role in influencing
plumage variation, raising the possibility that the plumage variation may be
more climate-driven than indicative of relationships. The genomic data don’t
even support the separation of pulchellus alone. A
modern analysis of geographic variation is needed to gain a better picture of
species limits in this complex.”
Comments
from Jaramillo:
“NO at this time, this is a messy situation and requires a more complete
sampling of vocal characters as well as genetics.”
Comments
from Gustavo Bravo (voting for Remsen): “NO. As everybody
has very well pointed out, the situation is messy, and we lack sufficient data
to determine species limits. To me, it is critical to assess gene flow across
potential contact zones between pulchellus and canadensis (e.g.,
Táchira depression), and the extent of plumage and vocal variation across the
whole complex."
Comments
from Bonaccorso:
“NO. Although pulchellus has has enough plumage characters to be diagnosable, a better
understanding of contact zones and genetics (more samples across the
distribution of all the subspecies) is necessary to make an informed decision.
Also, a formal vocal analysis would be a huge plus to a future proposal.”
Comments
from Lane:
“NO. Whereas I suspect that a more thorough study of this complex will uncover
the evidence necessary to support this proposed taxonomic change, the evidence
simply isn’t available right now.
“I am troubled by the eagerness of WGAC to
influence change when the cases are woefully under-sampled. In fact, I question
the point of the WGAC efforts completely: to try to get all the major world
checklists aligned is a fool’s errand. The authors of all these competing
checklists clearly don’t use the same criteria for determining species limits,
or they would all already match, and most would be superfluous… but they don’t
match, and different users use these lists depending on their own ideologies
and purposes. And that we in SACC are asked to vote on cases, when other
checklists have already made up their minds, suggests that we are still able to
disagree with those checklists, and thus will not fall into line with the other
WGAC members, correct? So…. What’s the point here? In most cases, the evidence
available simply isn’t sufficient to allow us to make an informed decision by
SACC’s usual standards, so either we abandon those standards, or we are forced
to settle for substandard or (more often) entirely incomplete evidence. This
may not matter to many checklist users (some or most of whom just want more
species to tick off), but it does mean that any student looking for a potential
project will simply assume that many of these poorly supported scenarios are
“already settled” when in fact they are very much not. To me, that is a harmful
side effect of jumping to conclusions without sufficient evidence.”