Proposal (992) to South
American Classification Committee
Treat Tyto furcata as
a separate species from Barn Owl Tyto alba
Background:
Two
recent proposals to NACC (2018-C-13 and 2022-B-6) have considered the
taxonomic placement of various taxa within the Barn Owl (Tyto alba)
complex. Comments on both NACC proposals and one submitted concurrently to SACC
(#908) raised concerns about
the lack of analysis of vocal differences among taxa.
Although
pointed out in comments under the previous proposals, we highlight here a
distinctive and prominent flight call associated with mate attraction that is
uttered by New World Barn Owls and is absent in Old World Barn Owls. Based on
this and concordant genetic data, we recommend adoption of New World Tyto
furcata as a separate species from the Old World taxa. Work that might
refine understanding of the Barn Owl complex both within the New World and
separately in the Old World is discussed along with what is known about vocal
and plumage differences. Genetic data presented in the previous proposals are
included for the sake of completeness.
The
cosmopolitan Barn Owl (Tyto alba) has a long and complex taxonomic
history, with the American, African, southeast Asian, Australian, and many
insular taxa being considered full species at various points. The current AOS
taxonomy (AOU 1998) is largely based on Peters (1940) who lumped many
previously recognized species under a cosmopolitan Tyto alba, with 34
then-recognized subspecies. When the AOU expanded coverage to include the West
Indies and Middle America, T. glaucops (previously subsumed under T. alba
by Peters 1940) was recognized as a separate species given its sympatry with T.
a. pratincola (AOU 1983). More recently, some authors have opted to
consider the American furcata clade and the southeast Asian + Australian
javanica clade as two species separate from the alba clade of
Europe and Africa (e.g., Gill et al. 2024). Additionally, three insular taxa from
the Macaronesian islands are occasionally elevated to species level (Robb
2015), as are some insular taxa in the Indian Ocean and Indonesia. Many of
these insular taxa are much darker than their mainland counterparts, including
some with dark facial disks. These are all outside our area but highlight that
species limits in the complex are highly dynamic, and that insular taxa
especially are treated as full species by some authors.
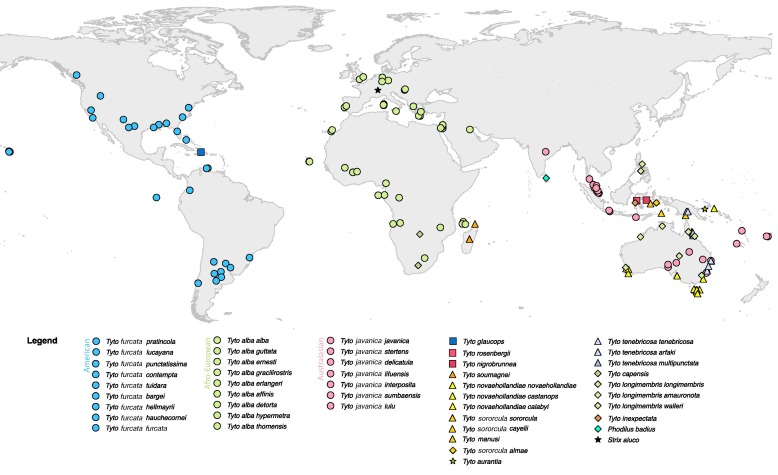
For
reference pertinent to this proposal, select taxa and subspecies groups
(based on Clements 2023) along with their respective distributions are listed
below:
• alba (Scopoli, 1769).
Subspecies group (4 taxa) Europe, n. Africa, and Middle East east to Iran
(hereafter alba ssp. group); the alba clade as a whole includes
the alba ssp. group plus six other subspecies that occur on islands off
Africa (5 taxa) and across sub-Saharan Africa (1 taxon, T. a. poensis),
each regarded as a separate subspecies group by Clements (2023).
• javanica (Gmelin, 1788).
Subspecies group (6 taxa) Pakistan east across s. Asia to Australia; also
referred to as javanica clade.
• furcata (Temminck, 1827). In sensu
stricto (s.s.) refers to T. a. furcata, a monotypic
subspecies group, White-winged Barn Owl (Clements 2023), of Cuba, Isle of
Pines, Cayman Islands, and Jamaica; elevated to species rank based on osteological differences by Suárez and Olson (2020);
sometimes regarded as part of tuidara subspecies group. For this
proposal, furcata clade or simply furcata refers to all 11
subspecies in the Americas, including tuidara group, currently
classified under T. alba (sensu lato, s.l.) and proposed
to be split as T. furcata.
• tuidara (J. E. Gray, 1827)*.
Subspecies group (6 taxa) ranges from Canada to Tierra del Fuego. Type locality
of tuidara is Brazil. [* see footnote on publication year]
• punctatissima (Gould & G. R.
Gray, 1838). Galápagos.
• pratincola (Bonaparte, 1838).
Mainland North America south to southern Mexico, recently to Hispaniola. Part
of the tuidara subspecies group.
• glaucops (Kaup, 1852)*.
Hispaniola. [* see footnote on publication year]
• insularis (Pelzeln, 1872). St.
Vincent south to Grenada. With nigrescens grouped as Lesser Antilles
Barn Owl (Clements 2023) or as a species (Suárez and Olson 2020); regarded as
subspecies of T. glaucops by Bruce (1999) and Gill et al. (2024).
• nigrescens (Lawrence, 1878).
Dominica. With insularis grouped as Lesser Antilles Barn Owl (Clements
2023) or as a subspecies under insularis (Suárez and Olson 2020);
regarded as subspecies of T. glaucops by Bruce (1999) and Gill et al.
(2024).
New
information:
Vocalizations:
One
of the primary issues raised by committee members in previous proposals is the
lack of analysis of vocalizations in the Barn Owl complex. Although no formal
analysis is yet published, we think that the qualitative analysis provided here
is sufficient to elevate the furcata clade to species rank. Across the
genus Tyto and within the Barn Owl complex there are a wide array of
both vocal and mechanical sounds. Here we focus on the context of vocalizations
associated with breeding, which is also the time when these owls are most
vocal. Two specific types of vocalizations are defined below: Screech
and kleak-kleak.
•
Screech: Categorized as either courtship or perennial (Robb 2015).
Recordings below are from https://soundapproach.co.uk/species/common-barn-owl/
(1) Courtship screech:
Used by males of alba clade (here specifically alba ssp. group), typically
given when perched but also in flight. Courtship screech in addition to
perched context is also longer and with shorter gaps between calls compared to
the perennial screech. Existence and context of this courtship screech is
unknown in the furcata clade (G. Vyn fide Robb 2015). Notably,
none of us has ever experienced a bird of the furcata clade screech from
a perch. This needs further investigation.
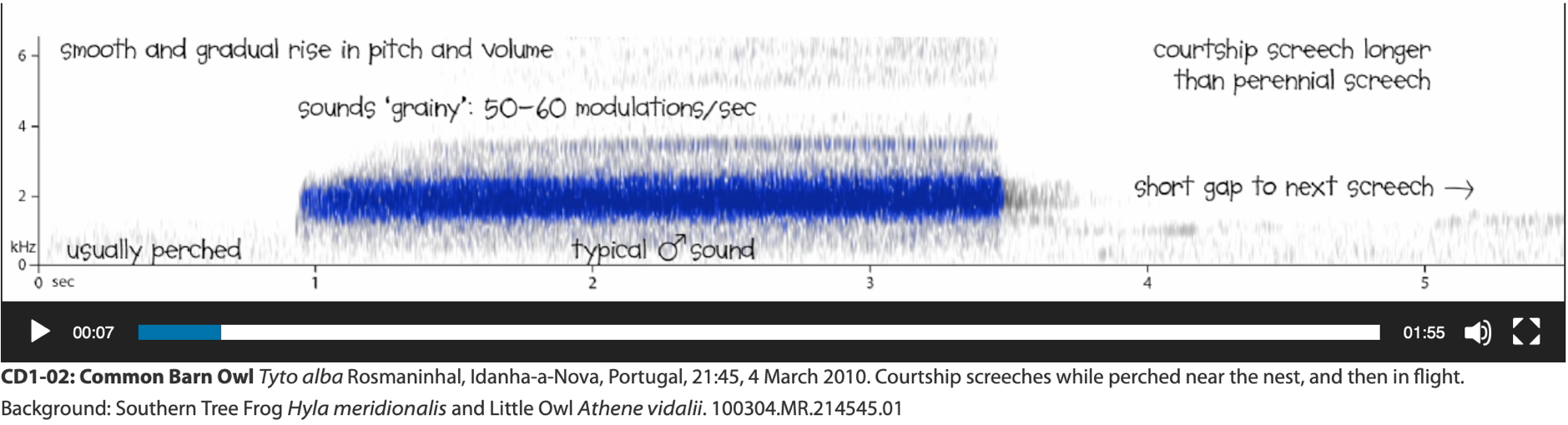
Spectrogram of courtship screech
by T. a. alba (Robb 2015)
(2) Perennial screech: Used by both sexes, uttered in
flight and less often from perch in alba clade but perhaps
never (or rarely?) given from perch in furcata clade. Further
investigation between the perennial screech of the alba clade and the flight calls of the furcata clade is needed, especially in the
context of whether the call is uttered when flying or perched.
• kleak-kleak (Vyn 2006): Given
in flight by furcata clade, perhaps most
often used by unpaired males (Gerrit Vyn pers comm fide M. Robb) or males in vicinity of
nest (Marti et al. 2020); presumed to have an important role in mate
attraction. Absent in both alba and javanica clades. Sometimes
categorized under terms like cackles, chirrups, or twitters.
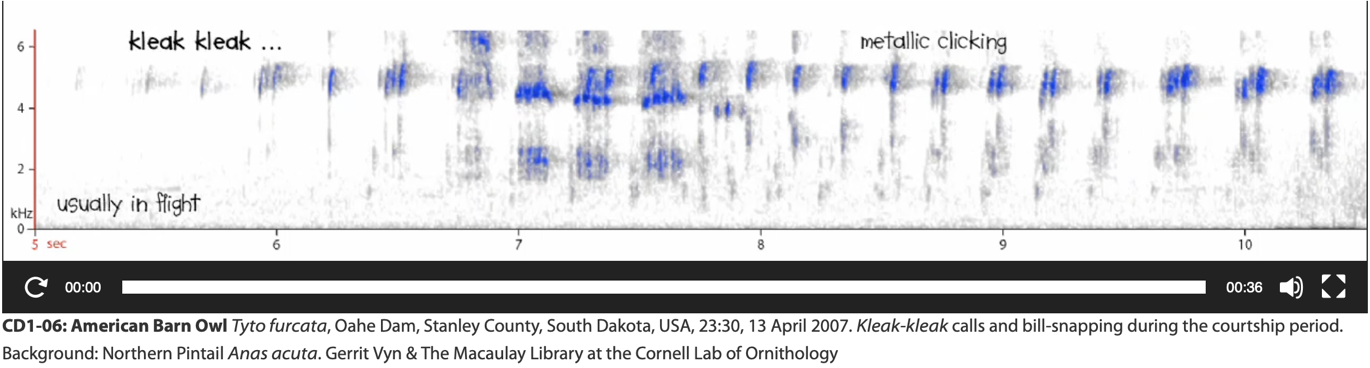
Spectrogram of kleak-kleak by T. a. pratincola (Vyn 2006)
The
screech (or scream in Marti et al. 2020) is the best-known
vocalization. The kleak-kleak call was described under “chirrups and
twitters” in Marti et al. (2020). We note that much published information
on vocalizations draws on Old World studies. Thus, it is important to heed the
warning in Marti et al. (2020):
“Other than anecdotal
notes, only unpublished information is available on vocalizations by the North
American race (E. McLean and B. Colvin pers. comm.). Some of the calls
described […] have not been positively documented for the North American race.”
Indeed,
much of the behavioral context and sounds ascribed to Barn Owls in the Americas
is adopted from Old World literature. Our summary here is guided in large part
by “The Sound Approach” (Robb
2015), with especially helpful material published by that author on Barn Owls
of the alba ssp. group here. One of us (O.J.)
perused the sonograms of all available Old World recordings on Xeno-canto
(1,080 alba clade and 62 javanica clade), plus a large selection in
the Macaulay Library. We found no examples
of kleak-kleak in either alba or javanica clades.
From
listening to recordings of many Tyto species, including glaucops
and various Masked/Grass owls it is clear that the loud screech call is fairly
conserved across the genus. There is some variation in length of the call among
species, and some have a whistled quality, but there is also much intra-taxon
variation in call length, perhaps related to whether these are courtship or
territorial, perennial screeches.
Typical
screech calls of the three clades are given below. For javanica and alba
clades, the screech tends to fade out and fall in pitch at the end of the
calls, unlike furcata clade which ends more abruptly and rises slightly
at the end:
alba: https://macaulaylibrary.org/asset/301733691
javanica: https://macaulaylibrary.org/asset/117266311 and https://macaulaylibrary.org/asset/271631421
furcata: https://macaulaylibrary.org/asset/50147
European
birds (alba) do tend to give longer screech calls than furcata,
while javanica are generally shorter but with a subtly different quality
than furcata. However, alba and javanica commonly have a
harsh whistled quality to the notes:
alba: https://macaulaylibrary.org/asset/235237551 and
https://macaulaylibrary.org/asset/367445881
javanica: https://macaulaylibrary.org/asset/273379781
Here
is an exceptionally long screech call from furcata: https://macaulaylibrary.org/asset/425012341
Non-screech
calls, when present, seem quite different among species. The Australian
Masked-Owl (T. novaehollandiae) utters a call
called a cackle that is said to be given in
courtship display flights by males circling over breeding territory (Higgins
1999, page 919). An example of that cackle call is here (https://macaulaylibrary.org/asset/539506871) and seems analogous
to the kleak-kleak call of furcata.
Likewise, analogous vocalizations exist in the two grass owls, T. capensis and T. longimembris
(Robb 2015).
The
kleak-kleak call of furcata is present across its range, with
recordings from California, Florida, and Brazil. Here are a few examples:
https://macaulaylibrary.org/asset/172455681
https://macaulaylibrary.org/asset/245778421
https://macaulaylibrary.org/asset/554918181
Critically,
this “kleak” call appears to be entirely absent
from both alba and javanica according
to The Sound Approach and our own perusal of
recordings. Robb (2015), quoting Gerrit Vyn (pers. comm.), says that “unpaired
males use this call most often…so it must have an important role in mate
attraction.” Marti et al. (2020) also report that males give the “kleak” call in the vicinity of the nest, soon after
leaving the daytime roost, and when approaching with food deliveries. Given
that analogous calls exist in T. novaehollandiae
and other Tyto, we suspect it has been lost in alba and javanica. Regardless, in our view this is a diagnostic vocal difference
between the clades.
In our personal experience, this “kleak” call is nearly always given in flight. For example, JLD recently witnessed (summer 2023) one bird giving the kleak-kleak call in fluttery flight almost nonstop for a few minutes as it circled a lit up area near a known nest. The only mention that we can find regarding the “kleak” call for alba is Bunn et al. (1982), who say that it is reportedly uncommon in Britain. This contradicts Robb (2015) who has extensive experience with the alba ssp. group in Portugal and elsewhere. Despite fairly exhaustive searches of databases online we are unable to find any recordings of this vocalization from anywhere in the Old World. This reference of the kleak call in Britain appears anecdotal and could refer to another call that Bunn (1977) called the kit-kit call.
We
feel it worth mentioning that no North American Field Guide or popular book on
owls, including König et al. (1999) and Weidensaul
(2015), mention the kleak-kleak call or its context in display. How did
the birding community miss this characteristic sound of New World birds? The
one source that does have it is Marti (1992), but none of us picked this up.
Genetics:
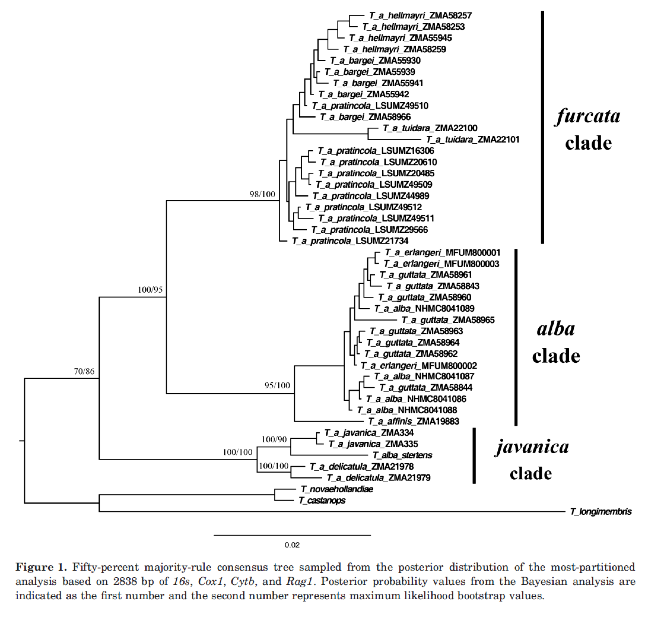
A
paper by Uva et al. (2018) analyzed two nuclear and five mitochondrial loci to
estimate a phylogeny of Tytonidae. This paper was included in the 2018-C-13
NACC proposal, and the proposal included a haplotype map that was based on a
single mitochondrial gene but did not include the phylogeny that was based on a
greater set of genes. That proposal did include the phylogeny from earlier work
by Alibadian et al. (2016) that was based on slightly fewer genes and many
fewer taxa. Although comments from many committee members considered the
genetic evidence inconclusive on its own, we include it in the current proposal
for the sake of completeness. Relevant figures from Alibadian et al. (2016) and
Uva et al. (2018) are reproduced below.
Phylogeny
from Alibadian et al. (2016):
Sampling
map, haplotype network, and phylogeny from Uva et al. (2018):
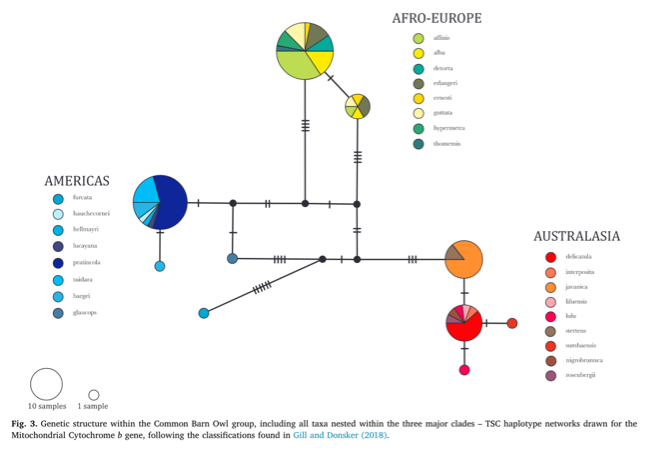
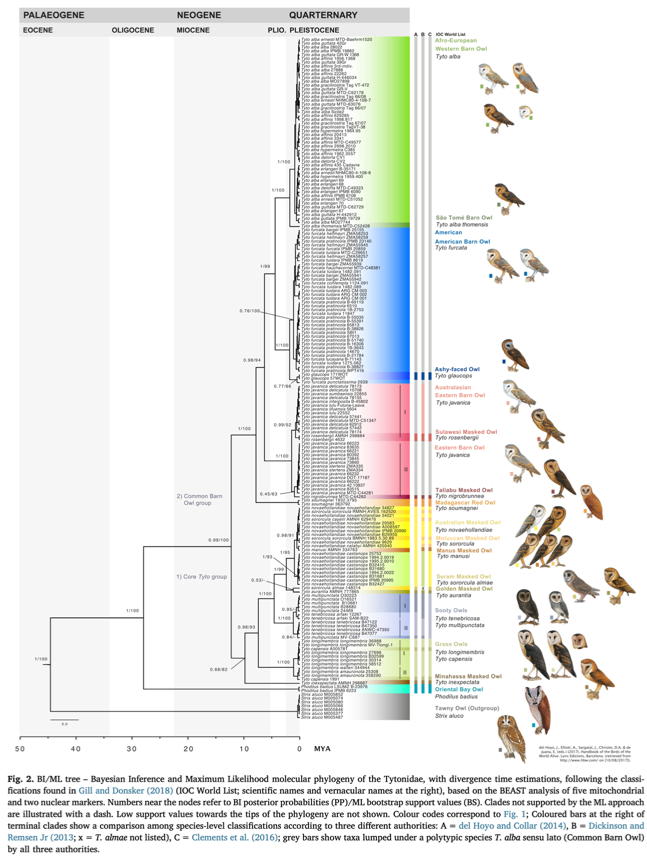
Based
on the genetic data, the current circumscription of Tyto alba comprises
three major clades: alba, furcata, and javanica, with the former
two being sisters. Uva et al. (2018) advocated elevating both furcata
and javanica to species rank. Whether the alba and javanica
clades should be treated as species is outside our purview and perhaps should
await potential future contact (see Additional Considerations, below).
A
few issues arise. First, Tyto glaucops is embedded within the furcata
clade, being sister to punctatissima of the Galápagos, the two in turn
being sister to the rest of the furcata clade. However, Uva et al.
(2018) note that “given the poor node support (0.77 PP/66 BS) putative genetic
distinctiveness of Caribbean and Pacific populations needs further
confirmation” and we agree with that assessment. Regardless, the species status
of punctatissima should be left to SACC.
Another
issue is that two dark taxa of the Lesser Antilles, nigrescens and insularis,
were not sampled by Uva et al. (2018); these taxa have been considered as
subspecies of Tyto glaucops or as their own polytypic species (Suárez
and Olson 2020). Given the lack of genetic and vocal information on these taxa,
we think it best to leave them as subspecies of alba (or furcata
if this proposal is adopted) for now, pending further study. Also see
Additional Considerations, below, regarding anecdotal information on Barn Owl
calls heard on Grenada where insularis occurs.
A
recent paper on Barn Owls of the West Indies by Suárez and Olson (2020) was the
basis for NACC proposal 2022-B-6, which did not pass but focused on the species
status of glaucops, nigrescens, and insularis plus some extinct
forms. Suárez and Olson (2020) analyzed osteological data from extinct and
extant Caribbean Tyto. They elevated the taxon T. a. furcata
Temminck 1827 of Cuba, the Cayman Islands, and Jamaica to species rank, leaving
tuidara J. E. Gray 1827 as the name for the American mainland species.
However, their osteological comparisons were to alba of Europe rather
than to pratincola of the United States, so the question of species rank
for furcata s.s. is unresolved. With regard to the priority of furcata
for American Barn Owls over tuidara if split from alba, see
footnote establishing that furcata has priority. Also note that furcata
s.s. is considered a separate subspecies group by Clements (2023) based
on the paler white plumage, especially of the wings. If, in the future, furcata
s.s. is elevated to species rank, then the name for the remaining
American barn owls would be tuidara Gray 1827. We note that Uva et al.
(2018) sampled one individual that they labeled as furcata s.s. (sample
number IPMB 20859), but no list of detailed sample localities is given in the
paper or supplementary data and there is no dot from Cuba, Jamaica, or the
Cayman Islands (the distribution of furcata) on their sampling map;
moreover, we do not recognize the museum acronym and were unable to find a
relevant record on VertNet or GBIF. Thus, it is unclear to us if true furcata
was sampled by Uva et al. (2018). Although it would be the nominate taxon of
the American clade, we think it extremely unlikely that it would be more
closely related to Old World taxa than to mainland North American taxa, so it
should not affect the separation of furcata clade from alba + javanica
clades. It may have implications for the taxonomy of other Caribbean Tyto,
however, if those are elevated to species rank in the future.
Plumage coloration:
Romano et al. (2019, Figure 2 reproduced here)
showed that plumage coloration appears closely tied to rainfall and
temperature. As can be seen in their maps, overall plumage coloration and spot
size are not drastically different between the three clades (furcata,
alba, and javanica).
Nevertheless, the plumage and size of several taxa within the Americas do
appear quite distinctive, e.g. punctatissima of
the Galápagos, bargei of Curaçao, and insularis/nigrescens of
the Lesser Antilles. Indeed, Ridgway (1914) separated these taxa and glaucops
from furcata s.s. and the remaining mainland American Barn Owl
taxa based on non-overlapping size, among other characters. Although not part
of this proposal, we would not be surprised if more detailed studies suggest
splitting more of these insular New World taxa. Interestingly, Ridgway (1914)
noted that bargei is similar to nominate alba of Europe in
coloration but is much smaller. We note that Uva et al. (2018) sampled bargei
and found it nested within the furcata clade.
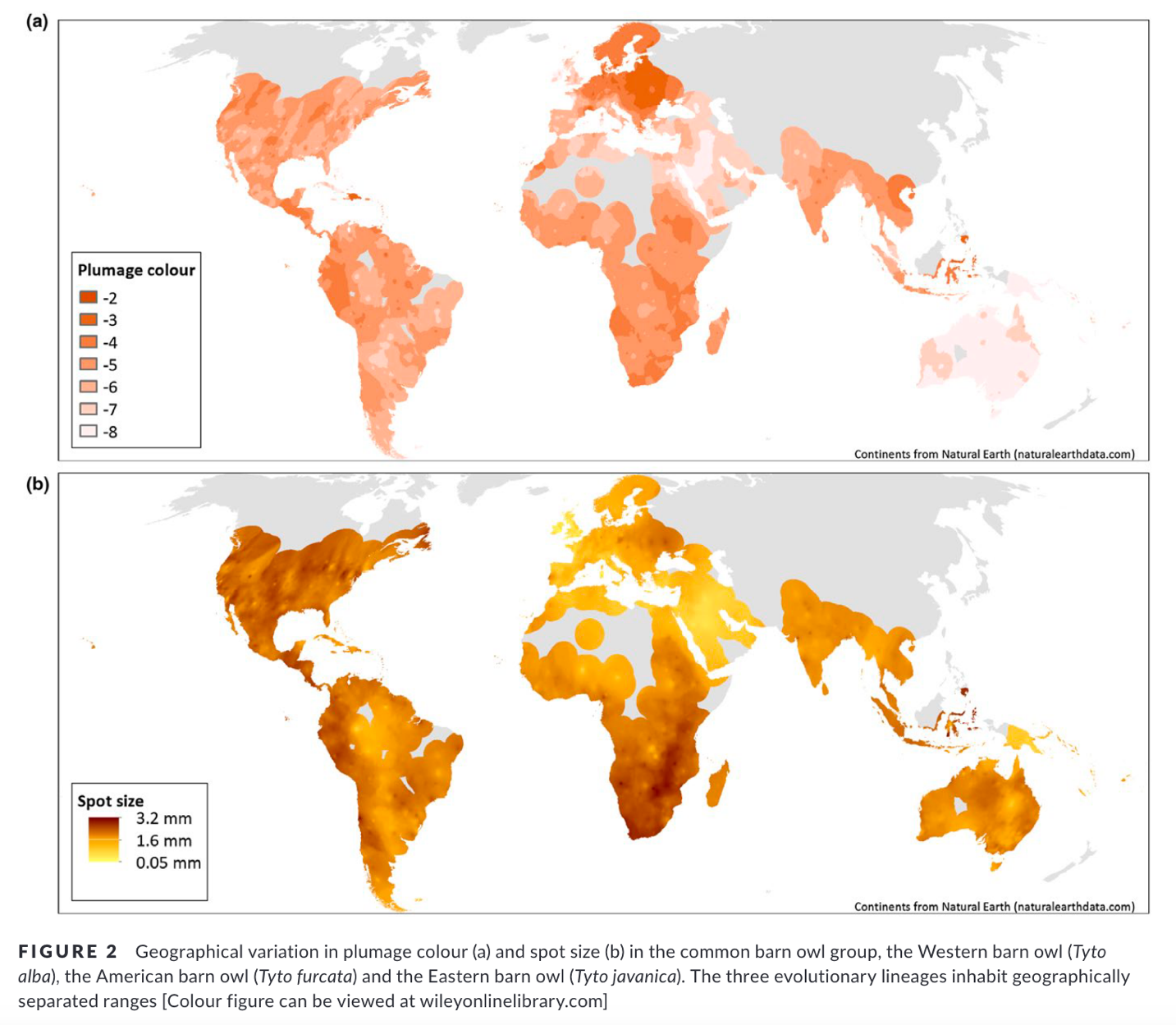
Additional
considerations:
We currently consider Tyto
glaucops unambiguously a separate species from T. alba s.l. based on
sympatry on Hispaniola. Earlier authors, however, considered glaucops
conspecific with alba s.l. (e.g., Hartert 1929, Peters 1940). On
Hispaniola, T. alba either colonized sometime after 1930 or was
overlooked before that (Keith et al. 2003). The source population is thought
likely to have been pratincola from the mainland or Bahamas (Marti et
al. 2020). Species limits considered by earlier authors were based on the same
issues that we are dealing with currently, namely plumage and vocalization
differences (but now also genetic) among allopatric insular populations.
However, once colonization by alba s.l. occurred, it became clear that glaucops
and alba s.l. were distinct species, a treatment followed ever since.
We
listened to available recordings of glaucops (of which there are few,
see examples below) and were not struck by major differences from furcata,
which raises additional questions. If furcata and glaucops are
sympatric, how are these being maintained as separate species despite the lack
of described vocal differences? The screech call of glaucops seems a bit
longer and more descending compared to furcata, which is interesting. If
that is the case, then there are some minor vocal differences in the
screech. A similar kleak call to the furcata clade is uttered by glaucops
and could be taken as further evidence that this is a major character
separating all New World barn owls (broadly speaking) from Old World barn owls.
This, then, would be more evidence for splitting furcata. On the other
hand, if plumage differences are keeping glaucops and furcata
separate, how does that fit into our understanding of species limits in the
genus given that plumage seems to covary with all sorts of things not related
to species limits? Maybe the fast evolution of plumage in the genus allows for
occasional evolution of drastically different-looking species?
Here
is an example of the cackling kleak call by glaucops:
https://macaulaylibrary.org/asset/175146681
And
recordings of its screech:
https://macaulaylibrary.org/asset/163149861
https://macaulaylibrary.org/asset/180725
We
note in passing that Alvaro Jaramillo suggested that Barn Owls Grenada (insularis)
gave vocalizations “much more like Ashy-faced Owl than Barn Owl” (Norton et al.
2005, page 512). Jaramillo’s analysis is repeated by Wiley (2021, page 209),
who himself reviews the taxonomic history of Barn Owls in the eastern Greater
Antilles through the Lesser Antilles. Note that east and south of glaucops
on Hispaniola (and formerly Puerto Rico; Suárez and Olson 2020), Barn Owls
occur on Dominica (nigrescens) and then on St. Vincent, some islands in
the Grenadines, and south to Grenada (insularis), with no confirmed
records for intervening Martinique and St. Lucia (Wiley 2021). To our ears, the
calls of T. glaucops do not sound that different from the furcata
clade so opining about calls of insularis on Grenada might be difficult
without careful analysis. Some recordings of insularis sound similar to
vocalizations of mainland furcata (https://macaulaylibrary.org/asset/541151851) but
others do sound quite different and rather like some recordings of alba s.s.
(https://macaulaylibrary.org/asset/317964701).
Given
that the node separating glaucops/punctatissima from furcata
is 1.75 Ma (Uva et al. 2018), it seems to us a reasonable yardstick extension
to consider the much older splits of alba and javanica as
different species from furcata (javanica vs. alba/furcata
is 6.25 Ma, alba vs. furcata is 4.35 Ma). The alternative here is
that glaucops/punctatissima are a recent offshoot from furcata that
(unambiguously in glaucops) evolved reproductive isolation, while furcata
and alba have not. We think that this is unlikely given that glaucops
seems to have evolved reproductive isolation despite limited or no differences
in vocalizations, while the limited vocal data we have indicate very distinct
vocalizations between furcata and alba + javanica (primarily the
lack of a “kleak” call in the latter as well as existence of courtship
screech in at least alba ssp. group in alba clade). We also note
that the node uniting glaucops and punctatissima has lower
support (0.77 posterior probability/66% bootstrap) than most other nodes in
that part of the tree, so the furcata clade may not be paraphyletic with
broader genomic sampling. The node separating glaucops and punctatissima
is 0.44 Ma. Uva et al. (2018) did not provide confidence intervals on these
node date estimates.
There
is also limited evidence that at least furcata and javanica are
reproductively isolated. Populations from each of those clades were introduced
onto Lord Howe Island to control rats: T. a. delicatula from the
Australian mainland in 1923, and T. a. pratincola from the San Diego Zoo
in 1927 (Hindwood 1940). Birds from these two taxa were not known to
interbreed, and this was taken as evidence that the two should not be
considered the same species (Bruce 1999). The only Barn Owl specimens collected
from Lord Howe are of the Australian population, and no Barn Owls are known to
have persisted past the mid 1980s (McAllan et al. 2004). It is presumed the
American birds died out soon after introduction. This contact between the javanica
and furcata clades could suggest that assortative mating was taking
place, but the period of sympatry was brief compared to the longer period of
sympatry between pratincola and T. glaucops on Hispaniola. We do
note that javanica is the more distant clade in the phylogeny and does
not provide direct evidence of species rank for the alba clade versus
the furcata clade. However, it does suggest that multiple species exist
within the cosmopolitan Barn Owl.
Finally, the International
Ornithologists’ Union Working Group on Avian Checklists (WGAC) has recently
split Barn Owl into three species, elevating the javanica and alba clades
in addition to furcata. Although recognizing two Old World species is
outside our purview, support for this is based on morphological differences (Dick Schodde fide
T. Chesser) and genetic evidence showing that the javanica and alba
clades are not sisters (Uva at al. 2018). It is important to note, however,
that Barn Owls have expanded east across much of Iran starting in the 1990s
(Osaei et al. 2007). Prior to this, when the species was rare in Iran,
specimens were ascribed to T. a. erlangeri of the alba
clade (Vaurie 1965). The easternmost record in Iran (subspecies unknown) is at
Bam, Kerman Province (Osaei et al. 2007), which is 900 kilometers (560 miles)
west of the western limit of T. a. stertens of the javanica
clade on the Indus Plain, eastern Pakistan. Thus, future contact between javanica
and alba is possible, and further research would help to elucidate
whether reproductive isolating mechanisms such as vocalizations exist to
maintain species-level differences. Nevertheless, we think it is worth
separately considering elevating javanica to species rank to align with
this global checklist.
Recommendation:
We
recommend splitting Tyto alba (Scopoli, 1769)
into at least two species to recognize the vocal and genetic distinctiveness of
New World taxa as American Barn Owl, Tyto furcata
(Temminck, 1827). If the javanica and alba clades are considered conspecific, then Common
Barn Owl is typically used for the Old World taxa.
English names: American Barn Owl is in wide usage
by authorities that split furcata from alba, and we recommend
that it be adopted. American Barn Owl was used by Ridgway (1914) for pratincola.
Because of the possibility of paraphyly with glaucops and various other
taxa embedded within javanica, we think that “Barn Owl” should not be
hyphenated unless there is interest in renaming glaucops to “Ashy-faced
Barn-Owl”. “American” in this context refers to the two continents on which
this species occurs.
If the javanica and alba
clades are retained as conspecific, then Common Barn Owl is typically used for
the Old World taxa. However, the IOC (Gill et al. 2024) recognizes javanica
and alba as Eastern Barn Owl and Western Barn Owl, respectively.
Clements (2023) uses Eastern Barn Owl, Eurasian Barn Owl, and American Barn Owl
for the subspecies groups. These English names are not ideal and potentially
misleading (e.g. “Eurasian” occurs in Africa, and “Eastern” and “Western” could
be confused with eastern and western North America). Therefore, consideration
or solicitation of alternative names for the Old World taxa is merited.
Acknowledgments
and Footnotes:
David
Donsker helped research publication dates for relevant taxa. Alan Peterson’s
Zoonomen.net website provided notes and insights on the publication dates of
original descriptions.
* tuidara (J. E. Gray, 1827): This name was published at
earliest 1 December 1827. Gill et al. (2024), among others, use 1828, whereas
Bruce (1999) and Peters (1940), for example, use 1829. The name Tuidara Owl
of John Edward Gray appeared in part 14 of Griffith’s Animal Kingdom, and this
part was published 1 December 1827 (see table here).
Temminck’s “Strix furcata”
was published 30 June 1827 in livraison 73, plate 432 of the “Nouveau
recueil de planches coloriées” (see table here
and would therefore have priority regardless of the confusing dates ascribed to
tuidara. The date of 1827 was
used for tuidara
by Suárez and Olson (2020), presumably based Cowan (1969). We use it here for
the same reason.
* glaucops
(Kaup, 1852): We found conflicting dates for this publication. The fourth
edition of Howard and Moore checklist (Dickinson and Remsen 2013), Bruce
(1999), and AOU (1998) use 1853. Gill et al. (2024), Peters (1940), and older
publications use 1852. Note that Murray Bruce later agreed 1852 is the correct
date (see notes here).
Literature
cited:
Alibadian M., N.
Alaei-Kakhki, O. Mirshamsi, V. Nijman, and A. Roulin. (2016). Phylogeny,
biogeography, and diversification of barn owls (Aves: Strigiformes). Biological
Journal of the Linnean Society 119, 904–918.
American Ornithologists' Union. (1983). Check-list of
North American birds. 6th ed. Am. Ornithol. Union, Washington, D.C.
American Ornithologists' Union. (1998). Check-list of North American birds. 7th edition. Washington, D.C.: American Ornithologists' Union.
Bruce, M.D. (1999).
Family Tytonidae (Barn-owls. In Handbook of the Birds of the World, Vol. 5),
pp. 34–75 (J. del Hoyo, A. Elliott, and J. Sargatal, Editors). Lynx Editions,
Barcelona.
Bunn, D. S. (1977).
[Letters] The voice of the Barn Owl. British Birds 70(4):171.
Clements,
J. F., P. C. Rasmussen, T. S. Schulenberg, M. J. Iliff, T. A. Fredericks, J. A.
Gerbracht, D. Lepage, A. Spencer, S. M. Billerman,
B. L. Sullivan, and C. L. Wood. (2023). The eBird/Clements checklist of Birds
of the World: v2023.
Cowan, C.F. (1969).
Notes on Griffith's Animal Kingdom of Cuvier (1824-1835). Journal of the
Society for the Bibliography of Natural History 5:137–140.
Gill, F., D. Donsker, and P. Rasmussen (Eds). (2024). IOC
World Bird List (v 14.1). doi 10.14344/IOC.ML.14.1
Hartert,
E. (1929). On various forms of the genus Tyto.
Novitates Zoologicae 35:93–104.
Higgins,
P., Ed. (1999). Handbook of Australian, New Zealand and Antarctic Birds. Volume
4. Parrots to Dollarbird. Oxford University Press, Melbourne, Australia.
Hindwood,
K. A. The birds of Lord Howe Island. Emu 40:1–86.
Keith, A. R., J. W. Wiley, S. C. Latta and J. A. Ottenwalder. (2003). The
Birds of Hispaniola: Haiti and the Dominican Republic. British Ornithologists’
Union Check-list No. 21. The Natural History Museum, Tring.
König,
C., F. Weick, and J.-H. Becking (1999). Owls. A Guide to the Owls of the World.
Pica Press, Robertsbridge, UK.
Marti, C. D. (1992). Barn Owl. In The Birds of North America, No. 1 (A. Poole, P. Stettenheim, and F. Gill, Eds.). The Academy of Natural Sciences, Philadelphia; The American Ornithologists' Union, Washington D.C. https://doi.org/10.2173/tbna.1.p
Marti, C. D., A. F.
Poole, L. R. Bevier, M.D. Bruce, D. A. Christie, G. M. Kirwan, and J. S. Marks
(2020). Barn Owl (Tyto alba), version 1.0. In
Birds of the World (S. M. Billerman, Editor). Cornell Lab of Ornithology,
Ithaca, NY, USA. https://doi.org/10.2173/bow.brnowl.01
McAllan,
I. A. W., B. R. Curtis, I. Hutton, and R. M. Cooper. (2004). The birds of the
Lord Howe Island group: a review of records. Australian Field Ornithology 21
(Supplement) 1–82.
Norton,
R. L., A. White, and A. Dobson. (2005). The Spring Migration: West Indies &
Bermuda Region. North American Birds
59(3):511–513.
Osaei, A., A.
Khaleghizadeh, and M. E. Sehhatisabet. (2007). Range extension of the Barn Owl Tyto
alba in Iran. Podoces 2(2):106–112.
Peters, J. L. (1940).
Check-list of the birds of the world. Volume IV. Harvard University Press,
Cambridge, Mass.
Ridgway, R. (1914).
Birds of North and Middle America. Bulletin of the United States National
Museum 50, part VI.
Robb,
M. & The Sound Approach. (2015). Undiscovered owls, A Sound Approach guide.
The Sound Approach. (web version available here:
https://soundapproach.co.uk/undiscovered-owls-web-book/)
Romano, A., R. Séchaud, A. H. Hirzel, and A. Roulin. (2019). Climate‐driven convergent evolution of plumage colour in a cosmopolitan bird. Global Ecology and Biogeography 28:496–507.
Suárez, W., and S. L.
Olson. (2020). Systematics and distribution of the living and fossil small barn
owls of the West Indies (Aves: Strigiformes). Zootaxa 4830:544–564.
https://doi.org/10.11646/zootaxa.4830.3.4
Uva V., M. Päckert, A.
Cibois, L. Fumagalli, and A. Roulin. (2018). Comprehensive molecular phylogeny
of barn owls and relatives (Family: Tytonidae), and their six major Pleistocene
radiations. Molecular Phylogenetics and Evolution 125 (2018):127–137. https://doi.org/10.1016/j.ympev.2018.03.013
Weidensaul, S. (2015).
Peterson Reference Guide to Owls of North America and the Caribbean. Houghton
Mifflin Harcourt, Boston.
Wiley,
J. W. (2021). The Birds of St Vincent, the Grenadines and Grenada. British
Ornithologists’
Club Checklist Series, volume 27.
Vaurie, C. 1965. The birds of the Palearctic fauna: a systematic
reference non-Passeriformes. H. F. & G. Whitherby, London.
Vyn, G. (2006). Voices of North American owls. 2 CD’s and booklet. Ithaca.
Louis Bevier, Carla
Cicero, Jon L. Dunn, Rosa Alicia Jiménez, and Oscar Johnson
February 2024
Comments
from Jaramillo:
“YES to this proposal. I have been listening to and recording
a few Barn Owls over my house recently, and had the same general thought. How
could we have missed that the clicking call is the important vocalization in
reproductively related display, not the screech. I have been listening to a
pair over my house that sometimes displays overhead and actually seem to have
an aerial "dance" of sorts where they cartwheel over each other,
although it is hard to see as it is pretty dark even with the town lights. But
the birds are clicking at each other, and sometimes one of them screeches, but
the clicking call is the key.
“Regarding Ashy-faced, Lesser Antillean and Galapagos birds. The
possible genetic relationship between Galapagos and Antillean birds is not
surprising to me. This pattern happens over and over again in multiple taxa.
There is a Galapagos-Caribbean connection that has not been studied. Galapagos
mockers are sister to Bahama Mockingbird, Galapagos finches are sister to the
mainly Caribbean closed nest tanager radiation (the "quits"), even
the flamingo is a connection. I worked with a Cuban spider expert who mentioned
that Cuban spiders had some relatives in Galapagos. The fact that this has not
been looked at more closely is interesting to me. I assume that there are some
islands that are gone that connected the two before the Panama land bridge
closed. In any case, I bet that in the future there is a confirmation of an
Antillean/Galapagos Barn Owl clade.”
Comments from Robbins: “YES. What an in-depth proposal!
Kudos to Bevier et al. for the extensive perspective on this. Their proposal along with Alvaro's comments
about the clicking call seem to underscore the importance of that vocalization:
present in New World and apparently absent in Old World taxa. That coupled with
the genetic data support recognizing New World furcata as a separate
species from Old World alba. I vote yes in support of this split.”
Comment from Andrew Spencer: “Pieplow
(2017) does mention the voc (which he calls "chitter") and does say
it can be given by pairs in courtship.”
Comments
from Niels Krabbe (voting for Elisa Bonaccorso): “YES. Back in the
late 1960s I banded a brood of three Barn Owl chicks and had the luck to have
all three recovered. One had flown 10 km SW, another 150 km N, and the third
over 100 km E. Even though adult Barn Owls can be among the most resident birds
and spend their entire life inside a single barn, young birds have a great
capacity for dispersal, so I was fine with its worldwide distribution. As much
as I hate to see another cosmopolitan go (there seems to be only Peregrine
left), I see from the extensive, primarily mitochondrial comparisons by Uva et
al. (2018) that the division of the Common Barn Owl group into three major
clades is well-supported. The absence of the "kleak-kleak" sound in
both alba and javanica clades and its presumed importance in mate
attraction, as mentioned above, indeed supports species rank for the furcata
clade. I was a bit surprised to find that several Xeno-Canto recordings of
screeches by furcata were of perched birds, but far the most were in
flight. Personally, I have only heard the short perennial screech, and only
from flying birds. I was flabbergasted, when I first experienced a bird flying
back and forth while giving the “kleak-kleak” call, which I had never heard in
Europe.
“The proposal is elaborate and thorough, and I
can find nothing to add. As stated, it is puzzling, that two Tyto
species with quite similar vocalizations are both widespread on Hispaniola
without interbreeding, and, also as stated, it would be interesting to know if
the dark forms nigrescens and insularis (Lesser Antilles), belong
with glaucops or furcata, but this is all outside the scope of
SACC. As also discussed above, the mitochondrial similarity between glaucops
and the Galapagos punctatissima has low support and needs to be
substantiated by in-depth studies.
“I vote YES for ranking American Barn Owl as a
species Tyto furcata.”
Comments
from Areta:
“YES. I was already convinced in https://www.museum.lsu.edu/~Remsen/SACCprop908.htm
that there was overwhelming evidence for this split, and some
unanswered questions on other Tyto. It is not
that the "kleak-kleak" went unnoticed to Neotropical
ornithologists, but it is the lack of it in the Old World that we (or I at
least) were not aware of. Nice proposal, very thorough and clear.”
Comments
from Stiles:
“YES. Genetics and morphology found previously to support this split, but a
thorough study of all available recordings now clearly demonstrates an
important difference in vocalizations that tips the burden of proof onto those
who favor maintaining the single-species status quo.”