Proposal (1000) to South
American Classification Committee
Treat Thamnophilus shumbae as
a separate species from T. bernardi
Note:: This is one of
several situations that the IOU Working Group on Avian Checklists has asked us
to review because published treatments of the species differ, and WGAC has to
pick one. BirdLife International (del
Hoyo & Collar 2014) now treat these two as separate species. I have no first-hand knowledge of this
situation, so my goal here is to lay out as many facts as possible that might
be of potential relevance just to get the discussion started.
Background: As
currently defined (e.g. Dickinson & Christidis 2014), Thamnophilus
(ex-Sakesphorus; see SACC 278) bernardi
consists of two subspecies that show a common pattern of taxon pairs (Tumbesian
+ Marañon):
• T. b.
bernardi Lesson, 1844: sw. Ecuador (Manabí) to nw. Peru (S to Ancash),
including crossing Andes into upper Marañon Valley in Cajamarca (Schulenberg et
al. 2007)
• T. b.
shumbae (Carriker, 1934): Marañon Valley
Here
is the terrific plate by Hilary Burn from Zimmer & Isler (2003), here
somewhat desecrated by my green line to show T. bernardi broadly defined. Two extremes of male plumage variation are
shown for bernardi. The female
shown for bernardi has a solid rufous crown, but Zimmer & Isler
noted that some have varying degrees of black in the crown – see also specimen
photos below.

Zimmer
& Isler synonymized two previously described subspecies under nominate bernardi
because their examination of specimens indicated that they were just points on
a cline: piurae Chapman, 1923 (type locality = “Piura”), and cajamarcae
(Hellmayr, 1917; type loc = Tembladera, Cajamarca, 1200 ft.”). The type locality for bernardi is near
the north end of its distribution: Guayaquil.
Another taxon, baroni Hartert and Goodson, 1917 (TL = “Yonan
River, northeast of Trujillo”), was treated as a synonym of cajamarcae
by Cory & Hellmayr (1924) and Zimmer (1933), almost certainly because the
type locality of cajamarcae, Tembladera, is on the Yonán River, and cajamarcae
has priority (barely). Cory &
Hellmayr (1924) and Zimmer (1933) both recognized piurae and cajamarcae. Note that shumbae was described after
publication of Zimmer (1933), who had very few specimens from the Marañon
region.
Here are a set of photographs of
specimens of males and females from LSUMNS:
Males (no bernardi
available)


Females


New information:
Schulenberg et al. (2007, Bird of
Peru) noted the song of shumbae differed from bernardi in being
“much faster with distinct introductory and terminal notes” but were evidently
reluctant to make any taxonomic recommendation.
Kevin Zimmer was evidently the first person to notice this (fide
Dan Lane), so I am looking forward to Kevin’s comments especially.
Del Hoyo & Collar (2016)
treated shumbae as a separate species (“Maranon Antshrike”) from bernardi
and other 4 subspecies. Del Hoyo &
Collar’s assessment using the Tobias et al. point scheme is as follows
(courtesy M. Iliff):
“Maranon:
https://birdsoftheworld.org/bow/historic/hbw/colant3/1.0/introduction
Hitherto treated as conspecific with T.
bernardi and placed in Sakesphorus (see T. bernardi), but
differs in its smaller size, notably shorter tail (effect size −3.3; score 2);
buff-tinged whitish vs pale tan-buff underparts in female (2); much reduced black
breast patch in male, so that malar and ear-coverts are black-speckled whitish
rather than black or blackish (2); darker, less rufous dorsal coloration in
male (1); and faster song, with greater pace (2) and shorter note length (2)
(1). Monotypic.”
The vocal differences are based on Boesman (2016), who quantified
substantial differences in pace and note length. Boesman wrote:
“We
can thus conclude that shumbae
has a loudsong which is delivered much faster (score 2-3) and which consists of
much shorter notes (score 2-3). If we include the somewhat faster songs of bernardi when singing in
duet, effect size drops somewhat. Nevertheless, difference still allows for a
score of 2+2 = 4.”
As previously noted, I’m a big fan of Boesman’s notes as catalysts
to guide more thorough research, but he pumped out 457 of these in a short time
and just did not have the time to get them into the shape that would be
required for peer review. In this case,
the geographic distribution of the recordings would have been of interest. No attempt was made to employ directly the
Isler-Whitney scoring system for antbird songs in terms of whether the
different best fit the species or subspecies category.
Some sample recordings:
• bernardi from Guayas,
Ecuador, by John V. Moore: https://xeno-canto.org/258338
• bernardi from Lambayeque,
Peru, by Andrew Spencer: https://xeno-canto.org/45729
• shumbae
from Jaén, Cajamarca, ca. 30 km from type locality of shumbae, by
Fabrice Schmitt: https://xeno-canto.org/543063
• shumbae from Bagua Chica,
Amazonas: https://xeno-canto.org/45701
The differences between the two song types as described by
Schulenberg et al. and Boesman et al. are fairly obvious (to me).
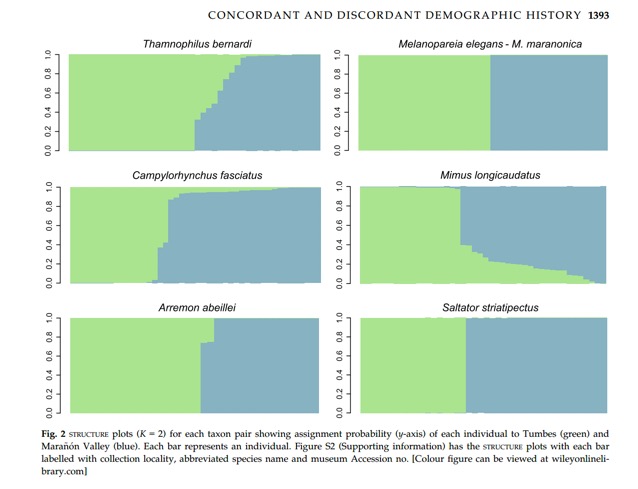
Oswald et al. (2017) included bernardi and shumbae
in their analysis of genetic divergence between Tumbesian and Marañon taxa in
six passerines. Using DNA sequencing and
SNPs, they found evidence for substantial gene flow from nominate bernardi
into Marañon shumbae. Although
this was the only one of their focal taxa known to be in direct contact, the
degree of gene flow was similar to or greater than that detected between
Tumbesian and Marañon subspecies of Mimus longicaudus and Campylorhynchus
fasciatus; in contrast, gene flow detected between one taxon pair (Melanopareia
elegans/M. maranonica) ranked as species was zero The other two subspecies
pairs showed zero (Saltator striatipectus peruvianus/S. m. immaculatus)
or negligible (Arremon a. abeillei/A. a. nigriceps) evidence of gene
flow (and those cases suggest a closer look is needed at their taxonomy.)
Harvey et al.(2020), with its
comprehensive taxon-sampling, included bernardi but not shumbae. The locality of their sample (ca. 50 km SE
Abra de Porculla; see voucher specimen by Dan Lane in specimen photographs)
places it in that part of the Marañon drainage within the range of bernardi,
in fact close to a putative contact zone.
Here is the relevant portion of the Harvey et al. tree. The green arrow points to Thamnophilus
bernardi and its sister species, a surprisingly closely related T.
atrinucha from Darién. The yellow
vertical line marks the depth in the tree of nodes at the species/subspecies
boundary according to current taxonomy.
Note that the branch lengths uniting bernardi and atrinucha
are actually shorter than those uniting almost any other traditional
subspecies. That’s a shocker. Boldly assuming that shumbae is more
closely related to bernardi than to atrinucha, then that branch
length would be even shorter. Although I
don’t think relative genetic distance data are useful in determining species
vs. subspecies rank, for those that do, note that shumbae would fall at
the extreme short end of branch lengths that unite subspecies.

Discussion
and recommendation:
All we really have to go on, in my opinion, is the difference between the
songs. I’m going to try to get an
antshrike song expert to take my vote on this.
If only there were some playback trials!
Obviously, it would be best if we could get a bird’s ear view of whether
the differences we can hear are sufficient to be a barrier to gene flow.
English
names: Del Hoyo and Collar (2016) used “Maranon
Antshrike” for shumbae and retained “Collared Antshrike” for bernardi. Because their ranges differ so much in size,
and because shumbae is more or less a peripheral isolate, retaining the
parental names is justified for bernardi. Marañon is not a good choice for shumbae
because Collared also occurs into the western Marañon drainage, in at least one
valley: the Río Chamaya (where the LSU sample is from). Also, we have a plethora of Marañon
Somethings. Do we really need yet
another one? I also foresee a problem
with birders naturally calling their first individuals as they enter the
Marañon drainage from the west “Marañon Antshrikes.” With these negatives, I think it’s worth a
separate proposal on English names.
Something like “Freckle-faced” would call attention to a conspicuous plumage
difference between the two and make people actually look at the bird. Dan Lane (pers. comm.) suggested Chinchipe Antshrike
or Pallid Antshrike. Jaen
Antshrike? Shumba Antshrike? Apparently, there is no current place called
Shumba but there is a Shumba Alto, a Shumba Baja, and a Cruz de Shumba all
north of Jaén. A local name might boost
conservation efforts in the region.
Further discussion postponed until if/when the current proposal.
References (see SACC Bibliography
for standard references)
Boesman, P. 2016. Notes
on the vocalizations of the Collared Antshrike (Sakesphorus bernardi). HBW Alive Ornithological Note 54: In Handbook
of the Birds of the World Alive. Lynx
Edicions.
OSWALD, J. A., I.
OVERCAST, W. M. MAUCK III, M. J. ANDERSEN, AND B. T. SMITH. 2017.
Isolation with asymmetric gene flow during the nonsynchronous divergence
of dry forest birds. Molecular Ecology 26: 1386–1400.
Van Remsen, June 2024
_____________________________________________________________________________________________
Vote tracking chart:
https://www.museum.lsu.edu/~Remsen/SACCPropChart968-1043.htm
Comments
from Lane:
“I'd be interested to hear what Kevin has to say, as I had heard that he and
the Islers had investigated the shumbae/bernardi situation nearly
20 years ago and the distinctiveness of the two was disappointingly less
obvious than they had hoped (or so I was led to believe), and so the project
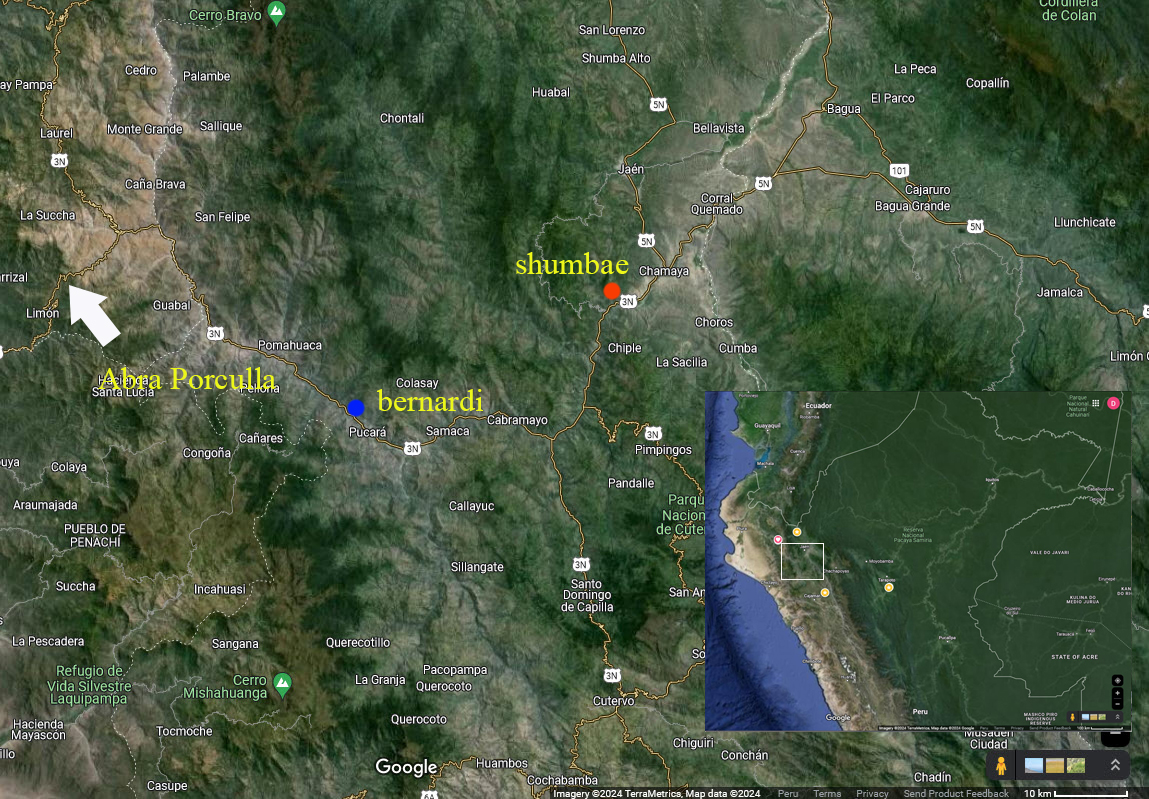
didn't progress further. My gut feeling is: given that both taxon groups are
found on the same side of the Porculla Pass (I have localities for the two that
are about 28 km apart, see map, straight-line-distance, along the Chamaya River
with no obvious biogeographic barriers between them, other than perhaps a
constriction of the valley walls along one stretch), and the distinctive female
plumage and voice, they probably would react to one another as different
species. But that's just a hunch and I've not been able either to document actual overlap or do much
playback experimenting. But if Kevin can elaborate on what he and the Islers
were able to ascertain of the two forms, that would help me decide how to vote
on this split.”
Comments
from Stiles:
“A tentative NO for recognizing T.
shumbae, but if information on different vocalizations can be
substantiated, this would tip the balance to a YES for me.”
Comments
from Jaramillo:
“NO – but a more substantial vocal analysis would be nice to have. It may tip
things the other direction into convincing all a split is the way forward….
Maybe?
Comments
from Jessica Oswald (voting for Remsen): “NO, based on the
evidence presented herein.
“Oswald et al. (2017) found that the best fitting demographic
model across all focal species was isolation with asymmetric migration from
Tumbes into the Maranon Valley. Estimates of divergence between
Thamnophilus bernardi subspecies
was during the Pleistocene (which could be as recent as ~12,000 years ago).
STRUCTURE recovered K=3 as the best K value (number of clusters/populations
represented in their dataset) for T.
bernardi. “The poorer fit of Thamnophilus [bernardi]
and Campylorhynchus [fasciatus] to a two-population (Tumbes – Maranon Valley)
model may be due to the shallow divergence of these taxon pairs, potentially
gene flow from an unsampled population in Tumbes, or genetic drift (Falush et
al. 2016).” Indeed, the geographic sampling of Oswald et al. (2017) of
T. bernardi
is restricted to a few sites in Peru so additional data from
Ecuador would be potentially informative.
“Further, I’m curious if there are any plumage or song differences
in individuals in Ecuador (coastal individuals or near Loja) as compared to
Peruvian birds.
“Playback data would be fantastic from across this species range.”
Comments from Areta: “I vote NO to the
recognition of T. shumbae. In our
WGAC exercise I have followed Harvey et al. in their interpretation that the
specimen sampled pertained to shumbae (and
this is what their supplementary table indicates), thus it was a surprise to
see that the specimen actually pertains to bernardi.
This does not change the interpretations substantially, as it is clear that a
study comparing shumbae, bernardi and atrinucha is needed. Oswald et al (2017) indicated that their
STRUCTURE analyses found K=3 to be the best. This is not shown in the main
STRUCTURE figure in the paper, but rather in the Figure S3:
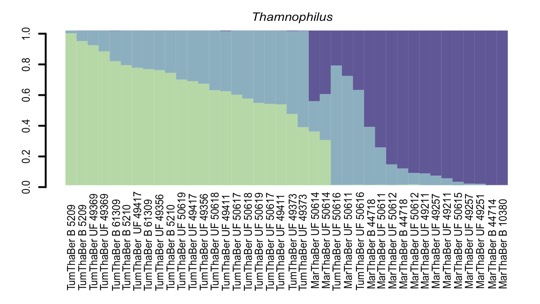
“Incidentally,
here is where a map would have been very useful to understand how geography and
genetics map together.
“The vocalizations sound indeed quite different, as
described in Schulenberg et al (2007), and Dan´s recordings of birds only 28km
apart provide a very suggestive piece of evidence. I see merit in the
hypothesis of shumbae
being a different species, but I cannot discard that the current subspecific
status is inadequate. This is the type of case in which a focus study can help
clear any doubts. It makes me uneasy to split taxa without a more clear
understanding of what seems to be, superficially, a good split, and the data on
asymmetric gene flow in Oswald et al. (2017) indicates that the situation is
not as clear-cut as it may seem in principle.”
Comments from Gustavo Bravo (voting for Elisa
Bonaccorso): “NO. As in the case of
S. canadensis (SACC 999), when you look at the
extremes of the range, there seems to be diagnosable phenotypic differences to
potentially elevate population to species status. However, as you move closer
to contact zones, things get messy and clinal variation associated with environmental
variation seems to be the case. What is missing is solid data. In T. shumbae’s
case, we at least have Jess’s genomic data, which seems to support my
interpretation.”
Comments from Zimmer: “YES. Of all of the many unfinished projects that
time conflicts have relegated to my backburner over the years, this one is one
of my bigger regrets. I made the first
audio recordings of shumbae on my first visit to the Marañon Valley back
in 2002 (I think). Finding it and
recording it was actually high on my radar, because Mort Isler and I were hard
at work on the antbird chapter for HBW Volume 8, and shumbae represented
something of a mystery, because there were no audio recordings of it in the
Islers’ extensive antbird vocal archives, and because the subspecies was
described (if I’m remembering correctly) from a female specimen, so we weren’t
even entirely sure about the parameters of male plumage characters. On this initial trip, I was actually scouting
out an itinerary for a fairly comprehensive Northern Peru tour, which had me
spending a lot of time in the Tumbes Region to the northwest, where other
recognized subspecies of S. bernardi were quite common. Accordingly, by the time I reached the
Marañon Valley, I had already seen, videotaped, and tape-recorded a number of bernardi. While searching for shumbae, I
periodically trolled with recordings of nominate bernardi I had just
made in the preceding week, and was frustrated to get zero response. Eventually, I stumbled across a spontaneously
singing/calling male shumbae and recorded it. With the bird in sight, I tried playback of
nominate bernardi songs and male-female duets (both of which I had
broadcast all over the area off-and-on over the preceding couple of hours),
neither of which elicited any detectable response from the male shumbae. After giving it a rest for a couple of
minutes, during which the bird continued to forage quietly, I then played back
the bird’s own spontaneously given vocalizations that I had recorded several
minutes earlier. This elicited an
immediate response, as the male approached me, raising and lowering its crest,
and resuming calling, before starting to sing.
I immediately began taping the song, and before long, a female appeared
and joined the male in singing and calling.
I was blown away by how much faster the song was compared to all of the
nominate bernardi that I had been hearing and recording for days. Over the next few days, and again, when I
returned in 2003, I made efforts to locate and tape record as many shumbae
as possible, and each time I found a new territory, I made a point of
performing similar playback trials, presenting the birds first with playback of
songs, calls and duets of nominate bernardi, followed by a break, and
then presenting them with songs, calls & duets of shumbae. In each instance, there was a complete
absence of response to playback of bernardi vocalizations, followed by a
strong and immediate response to playback of shumbae vocalizations. In the process of my investigations, one of
the things I was curious about, was establishing whether or not there was a
contact zone between bernardi and shumbae, so I worked my way
from the valley floor back upslope toward Abra Porculla, surveying for antshrikes
of either taxon as I went. I ended up
finding several more pairs of each, always assortatively mated, no sign of
intermediate phenotypes (in voice or plumage), and no actual contact zone. I conducted reciprocal playback experiments
with all of the bernardi that I found, presenting them with songs &
duets of shumbae, followed by a break, and then, songs and duets of bernardi. The results were similar – no bernardi
ever responded to songs/duets of shumbae, but did respond strongly to
songs of other bernardi. I did
find territories of bernardi and shumbae that were separated from
one another by as little as 30-35 road km, with no obvious barrier to dispersal
(e.g. large river, steep canyon, major habitat-shift other than gradually
increasing aridity the lower downslope you went toward the Marañon Valley floor). It seemed to me that the two taxa were
essentially parapatrically distributed in this area, and that they were
behaving as would be expected from two distinct species under the BSC.
“Needless to say, I was pretty
pumped about this, and so was Mort Isler when I returned home from the first
trip and told him about it. We
immediately got to work on it, and Mort began a thorough vocal analysis comparing
of all of my shumbae recordings, with what was already a pretty
extensive set of archived bernardi recordings from throughout its range.
“Somewhere during this process (I
can’t remember if it was after my first trip or after my second, the following
year.), Dan Lane returned from a Peru trip, wherein he had independently found
and recorded shumbae for the first time.
Dan also recognized the distinctiveness of shumbae, and contacted
Mort, who told him we were already actively working on it. My recollection is that Dan contributed
additional recordings and data, and, at Mort’s request, made detailed
descriptions of specimens of shumbae from the LSUMNS collection, and it
was our intention to add Dan as a coauthor on the paper.
“For the vocal analysis, Mort
divided the overall sample of S. bernardi (sensu lato) recordings into 5
clusters, 4 of which corresponded to the named subspecies at that time (bernardi,
piurae, cajamarcae, and shumbae) along with a 5th cluster
of recordings that fell midway between the type localities of piurae and
cajamarcae, and which Mort labeled as piurae/cajamarcae for
convenience, with no taxonomic implications.
This provided a basis for examining the possibility of clinality in
vocalizations. Mort analyzed recordings
of 105 individuals from 32 different localities. Although all vocalizations were examined
spectrographically (e.g. for note shape differences), only loudsongs were
measured. Whenever possible, Mort
measured 3 songs per individual. He
obtained 63 measures and ratios reflecting 12 vocal characters, including the
number of notes; duration, pace (notes per second), change in pace, note shape,
change in note shape, note length, change in note length, interval (space)
length, change in interval length, frequencies of note peaks, and change of
note peak frequencies.
“Recording measurements were
analyzed in three steps. First, loudsong
measurements for males and females were compared within each population. Second, data for both sexes were aggregated
by population, and characteristics of loudsongs of the five populations were
compared. Third, data for shumbae
were compared to an aggregate of the remaining populations. Spectrographic examination allowed us to
identify 6 basic types of vocalizations:
loudsongs, softsongs, bark, bark-roll, growl, and caw (caws were highly
variable in length and pitch). Except
for loudsongs, no diagnostic differences could be distinguished between
populations for any of these types of vocalizations, although it should be
noted that the inventory of calls was very small for some populations. The remainder of the analysis, and all
measurements, was confined to loudsongs.
‘The first step, a comparison of
male & female loudsongs within each population, found no diagnostic sexual
differences, although our sample contained no recordings of female loudsongs
for cajamarcae, and only 2 for nominate bernardi. Consequently, male and female recordings were
combined for the remainder of the analysis.
“In the second step, the combined
date for both sexes were compared for the five populations. Diagnostic differences were found between shumbae
and the remaining 4 populations in pace and change of pace (degree of
acceleration), with the acceleration in shumbae songs being most
pronounced in the second half of the song.
There were no diagnostic vocal characters distinguishing any of the
other 4 populations from one another.
“In the final step, data from shumbae
were compared to the combined values for all of the (aggregated) remaining 4
populations. Again, loudsong differences
between shumbae and the aggregated populations were found to differ
diagnostically in both pace and acceleration of pace.”
“Because no vocal characters were
found to distinguish loudsongs of populations described as nominate bernardi,
piurae, and cajamarcae, and because our examination of specimens
revealed that alleged plumage distinctions between the named subspecies were
inconsistent, and showed evidence of clinality, we ended up merging cajamarcae
and piurae with nominate bernardi, in our HBW chapter (2003),
wherein we recognized only two subspecies, bernardi and shumbae. On the other hand, our vocal analysis
revealed that loudsongs of shumbae were distinguished from the remaining
populations by two independent vocal characters. Although fewer in number than typically found
in syntopic pairs of closely related thamnophilid species (Isler et al 1998),
the two independent diagnostic vocal characters, combined with the finding of
parapatry, strongly indicated to us, that shumbae and bernardi
should be considered specifically distinct.
“We then asked Robb Brumfield to analyze tissue samples of bernardi
and shumbae (I don’t remember if this was a bird that Dan collected – he
could probably fill that part in.), to see if molecular distance coincided with
the vocal differences. In piecing
together old email threads, I see one from Robb to Mort, dated 6/24/2004 that
said “The two Sakesphorus bernardi bernardi and S. b.
shumbae are divergent by only 0.6%.
That’s pretty minimal for species.”
That was followed 2 days later, by an email from Mort to me, in which
Mort relayed to me the following: “I
talked to Robb after I spoke with you yesterday. He is quite open to the possibility that
molecular distances found under current procedures might not correspond to
species level differences under the BSC, and he agrees that the paper should go
ahead.”
“I remember that we were a little
discouraged that the molecular data were not strongly supportive of our
taxonomic conclusions, but also thinking that perhaps the divergence was just
really recent in evolutionary terms, and the fact that the two forms seemed to
be existing in parapatry, with no phenotypic signs of intermediacy in plumage
or vocalizations, meant that they were behaving as distinct biological
species. And, while it was true that the
vocal analysis only revealed 2 independent, diagnostic vocal characters (as
opposed to the 3-character yardstick of Isler et al, 1998), the ranges of those
two characters were not only non-overlapping between shumbae and bernardi,
but mean values were not even close statistically.
“Somehow, although it was our
intent to proceed with the paper and make the case for separate species status
for shumbae, we lost momentum.
Mort was concurrently involved in a big paper that was nearing
completion, and I got consumed with work related stuff. I think I was also hoping to make another
trip back to northern Peru to try to further investigate the contact zone
before we submitted the paper, and when that trip didn’t come off, we
completely lost all momentum, and we were both absorbed in a series of other
projects that always seemed to take precedence.
Now, so many of the details seem like ancient history, and it would take
some effort to recreate everything.
“It should be kept in mind that
although we did not identify any diagnostic differences in calls between shumbae
and the other four populations of bernardi, that sample sizes of calls
were limited, and inspection of calls was limited to visual inspection of
spectrograms, without any measurements being made. There are also other groups of closely
related thamnophilid antbirds which are known to differ in their loudsongs, but
which do not differ appreciably in their calls.
Although we identified only two diagnosable vocal characters, the extent
of the differences in those two characters were striking, and the potential
significance of them as isolating mechanisms is reinforced by my series of
reciprocal playback trials. Also, I
think it bears pointing out that our vocal analysis did not find any diagnostic
differences in vocal characters over the much broader range of bernardi,
and then, over the space of < 30 km, with no intervening geographic barrier,
bernardi is replaced by a taxon that differs in morphometrics, plumage,
and at least 2 vocal characters.
Regardless of what the genetics are suggesting, to me, that’s a pretty
strong case for treating these two taxa as separate species.”
Comments from Remsen: “I now recommend
a YES because Kevin’s comments above actually fill in the blanks perfectly in
the information gaps I identified in the proposal. I am especially impressed by the first-hand
account of the playback trials, and I am highly influenced by the documented
abrupt turnover from bernardi to shumbae. Parapatry without evidence of gene flow is
THE strongest evidence one can get for species rank in non-sympatric taxa. As for genetic distances … well, as you can
anticipate, I don’t put much stock in them – they measure time-since-separation
divergence at neutral loci, which is correlated broadly with speciation, but
the correlation is so weak that I never think it should be used as primary
evidence except at its extremes. If one
could measure just the “genetic distance” between the gene complexes that
control song in this case, I suspect that the degree of divergence at those
loci would be substantial, comparatively speaking. These taxa may not have been separated for
very long but obviously long enough for songs to diverge to the point of not
recognizing each other as conspecifics.
“The only reason I can see now, in
my opinion, for rejecting the proposal is that the critical information is not
published in a journal. I can see that
side of the argument, also.”
Additional comments from Bravo: “I read Kevin’s vote –
some bits of which I already knew – and I think it is very telling. However, I
stick to my NO vote.
“It is still necessary to see all those data in published form and
explicitly test for clinal variation. I spent the Summer of 2017 following shumbae
around in the core of its range and the Summer of 2019 following bernardi
in Tumbes. As I said, those two extremes are decisively different vocally
(loudsongs), morphometrically, and in plumage. Still, I would like to see how
those traits vary across the entire range of the species, particularly given
the low genetic divergence and the strong signal of admixed individuals
presented by Oswald et al. (2017). What I think is missing from Kevin’s
described vocal analyses is an explicit test of gradual variation across space.”
Comments
from Lane:
“YES. Wow, well Kevin really delivered there! I should add my own addendum to
his tale. He and I overlapped at the start of a tour he ran in 2002, in which
Kevin kindly allowed me to join him and his group on a day trip to Lomas de
Lachay. During that day out, he told me about his discovery of the different
voices of the two T. bernardi, and that he had found coastal type birds
east of Abra Porculla, which was what inspired me to try to collect some when I
returned to the area in 2003, and I successfully got one female (which,
apparently, is the one that was used in the Harvey et al tree, but was
erroneously identified as shumbae). I had collected a series of T. b.
shumbae in Sept 2002, with recordings, and those tissues should be
available at LSU.
“Anyway,
given the work Kevin has done in playback experiments, the fact that we both
were able to document the two taxa along the same river valley within ~35 road
km of one another without evidence of interbreeding, and the fact that the
coastal (nominate) group of T. bernardi shows no clinal or other shift
in song characters over a huge region, I think the evidence is strong, if
unpublished, in support of species status for T. shumbae. YES.”
Additional
comments from Areta:
“NO. The genetic data is suggesting that something
complex can be happening here, and like Gustavo, I would like to see a proper
paper analyzing this. If there is a mismatch between genomics and phenotype,
then it also should be explained. I am impressed by the amount of work that
Kevin has done, and taking a formal look at the vocal and plumage data in
association to genetics should clarify what´s happening here. Sometime ago I
tried to lure Peruvian students into this complex, but apparently failed to
create enough enthusiasm. This proposal is now making the case very appealing
for further study.”
Additional comments from Robbins: “Reading all those great
comments/data by Kevin leaves little doubt that shumbae should be
considered a species. So, I change my vote to YES.”
Comments from Claramunt: “NO. Kevin’s observations are
very suggestive of reproductive isolation, but the genetic data suggest free
gene flow between the two taxa. Evidently there is a lot of geographic
variation in bernardi
(plumage and voice) that needs to be analyzed and in general, we
need more published information to make an informed decision.”
Additional
comments from Stiles:
“YES. Although the data are clearly not
all there yet, I think that the detailed field experience of Kevin,
especially including the playback experiments, may have tipped the balance
toward considering shumbae a distinct species – so, a tentative YES
here.”
Comments from Rafael Lima: “I was recently reviewing the results of
Oswald et al. (2016) and Boesman (2016) for a paper on antbird speciation, and
I wanted to share some thoughts in the interest of understanding the rationale
behind the votes in favor of splitting Thamnophilus bernardi.
“The song differences between bernardi and shumbae are
indeed impressive, greater than those between several closely related syntopic
antbird species. Most importantly, the fact that they strongly discriminate
each other's songs suggests that these differences could generate strong
premating isolation. However, the genetic evidence suggests substantial gene
flow between them.
“This case initially reminded me of the situation with Hypocnemis
and Willisornis, where vocally distinct antbirds hybridize despite
marked vocal differences (Pulido-Santacruz et al. 2018, Proc. B. 285: 20172081;
Cronemberger et al. 2020, Evolution 74: 2512-2525). I wondered whether
postzygotic isolation might be at play, as seen in those genera. But this seems
not to be the case with bernardi and shumbae. For instance,
consider samples FLMNH 50616 and LSUMZ B-44718 in Fig. S2 of Oswald et al.
(2016). Both exhibit noticeably admixed genotypes. According to Supporting
Information Table S1 of Oswald et al. (2016), sample FLMNH 50616 is from
"Coto de Caza El Angolo, Los Pilares, 4.5 km SE Fernandez (-4.22,
-80.84)" and sample LSUMZ B-44718 is from "Las Juntas, junction of
rios Tabacomas and Chinchipe (-5.62, -78.53)." These two locations are 300
km apart. It therefore appears that genetic admixture is taking place over 300
km between bernardi and shumbae.
At k = 3, admixture seems even more extensive (see Fig. S3). Unlike Hypocnemis
and Willisornis species, which form very narrow and strongly bimodal
hybrid zones, bernardi and shumbae appear to form a wide zone of
genetic intergradation.
“One possible interpretation is that assortative mating between bernardi
and shumbae may be strong but incomplete. Recent theoretical work
indicates that assortative mating effectively maintains species boundaries only
when it results in essentially complete premating isolation (Irwin 2020, Am.
Nat. 195: 150-167). If assortative mating is strong but incomplete, it may not
generate strong overall reproductive isolation unless accompanied by postmating
isolating barriers. It is plausible that bernardi and shumbae
exhibit strong but incomplete assortative mating based on vocalizations and
weak or nonexistent postmating barriers, resulting in weak overall reproductive
isolation.
“My interpretation is that bernardi and shumbae are
experiencing extensive gene flow *despite* marked vocal differences and strong
behavioral song discrimination. Since overall reproductive isolation is what
determines species limits, the currently available evidence suggests a single
species. I am curious to understand the rationales behind the "YES"
votes, given the evidence of extensive gene flow.”
Response from Remsen: My
response to Rafael is that not all gene flow is alike. The evidence for extensive gene flow in this
case comes from SNP data, i.e., differences in which a single nucleotide in a
DNA sequence is replaced by another.
Whether these have any relevance to the bird’s biology and whether these
are influenced directly by natural section is not known. As known for a long time (e.g. in Brumfield Manacus
papers), presumably neutral genetic differences (e.g. mtDNA sequence
differences) are much more labile in terms of gene flow, whereas phenotypic
characters likely under strong selection, e.g. things important to reproductive
isolation such as song and plumage, do not necessarily show the same pattern
and may show abrupt shifts from one taxon to the other. My interpretation of both the vocal and
plumage data in this system is that there is an abrupt shift, which suggests
assortative mating at contact, albeit incomplete.”
Comments from Bonaccorso: “My vote
on this one was given to Gustavo and I agree with him that without a formal
clinal analysis, Kevin’s ideas remain hypotheses that need to be tested.
Therefore, I support his NO vote.”
Comments from Jaramillo: “YES. I
am a big fan of playback experiments to ask the birds what they think. Even a
small number of experiments on suboscines can do the trick. The most
informative result is that the vocalization is ignored, while same taxon
vocalizations are not ignored. Yes, it would be great to have this published,
but enough is out there and with these additional comments I am more
comfortable with a YES vote at this point.”