Proposal (1047) to South American Classification Committee
Revise Turdus assimilis/T.
albicollis complex as consisting of four species: (A) treat daguae
as a separate species from T. assimilis, and (B) treat the phaeopygus
group as a separate species from T. albicollis
Note
from Remsen:
This
proposal was written for NACC to treat the T. assimilis-T. albicollis
complex as more than two species, with an emphasis on daguae because it
occurs in the NACC area. Note that we
dealt with this issue in 2021 – see SACC proposal 922. Oscar, Jacob, and I have modified this
slightly to include the option of also recognizing Turdus phaeopygus
as separate from our T. albicollis, despite this not being the main
thrust of the NACC proposal, in case we consider the information herein as
sufficient to treat the phaeopygus group as a separate species. If rejected, we could potentially reconsider
that in a separate, more detailed proposal that focuses strictly on phaeopygus. Currently, SACC treats daguae as a
subspecies of otherwise Middle American T. assimilis and phaeopygus
as a subspecies of South American T. albicollis.
Effect
on SACC:
Splitting T. daguae and T. phaeopygus from T. assimilis
would result in two additional species for the SACC area.
Description of the
problem:
The
assimilis/albicollis complex is found
throughout much of Middle and South America and comprises four main subspecies
groups. The assimilis group is found
in the highlands from northern Mexico in all mountainous regions south to
central Panama. The taxon daguae is
currently considered a subspecies of assimilis
and is found in the lower foothills of the Chocó biogeographic region, from
southwestern Ecuador north to eastern Darién. In the Amazon Basin and the
Guiana Shield, three subspecies represent the phaeopygus group of T.
albicollis. These reach as far north/west as the Sierra de Santa Marta and
as far south as northern Bolivia, Mato Grosso, and Pará. The nominate albicollis group is found in
southeastern Brazil, Argentina, and Uruguay, with an isolated population
(subspecies contemptus) in the Andes
of Bolivia, in close proximity to lowland phaeopygus.
Taxonomic history
The
taxonomic history was covered thoroughly in NACC proposal 2022-A-4 / SACC
proposal 922. Briefly, daguae was
described as a species in 1897 by Berlepsch, but of course in a different era
and different species concept. Peters (1964) considered all four groups to be
members of one species, T. albicollis.
Most authorities have considered daguae
as a subspecies of T. assimilis (of
Middle America) based on plumage similarity. A few recent authorities (e.g.,
HBW BirdLife) have considered daguae
as a subspecies of T. albicollis (of
South America) based on vocal similarity or more recently as a separate species
(Ridgely & Greenfield 2001).
In
early 2021, the Working Group Avian Checklists (WGAC) addressed the placement
of daguae and voted 6-0 to consider daguae as a species separate from both T. assimilis and T. albicollis. However, two WGAC committee members noted that they
wanted to hear from NACC and SACC and would reconsider their votes if those
committees disagreed with that conclusion. Later in 2021, Van Remsen submitted
a proposal to split daguae
concurrently to both NACC (proposal #2022-A-4) and SACC (proposal #922). Those
proposals both failed (respective votes: 4-7 and 4-4), with both committees
retaining daguae as a subspecies of T. assimilis. WGAC has not reconsidered
its vote since these NACC and SACC proposals.
The
SACC proposal is here: https://www.museum.lsu.edu/~Remsen/SACCprop922.htm
And
the NACC proposal (and comments) are here: https://americanornithology.org/about/committees/nacc/current-prior-proposals/2022-proposals/
The
WGAC proposal was based on the same information as what was contained in the
NACC and SACC proposals. We encourage the committee members to read the
NACC/SACC proposal, and especially the comments by committee members on both
proposals.
New information:
No
new information since 2021. We are here again addressing this issue in advance
of the publication of the WGAC checklist, to attempt to minimize discrepancies
between NACC and WGAC. Committee members in both NACC and SACC who voted
against changes to taxonomy raised two main issues: 1) genetic sampling was
insufficient, both spatially and in number of loci, and 2) no formal analysis
of vocalizations was conducted. Although both issues could use additional
research, we present additional clarification on both, that together clarify
some of the issues raised by committee members.
Regarding
genetic sampling, most data come from the mitochondrial tree presented by
Núñez-Zapata et al. (2016) and was included in NACC 2022-A-4 / SACC 922. That
tree showed a sister relationship but with a deep divergence between daguae and assimilis. All the samples of daguae
came from southern Ecuador, far from any potential contact zone, but note that
based on distribution we do not believe that these taxa come into contact. SACC
committee members also noted that there may be multiple species within T. albicollis, and that Núñez-Zapata et
al. (2016) sampled only the nominate subspecies group of southeastern Brazil,
Argentina, Uruguay, and Andean Bolivia. We checked this to be certain, as both
subspecies groups occur in Bolivia, and based on the sampling localities in
Núñez-Zapata et al. (2016), confirmed that both Bolivian samples were from
Andean localities, so represent contemptus
of the albicollis group. We can
confirm, therefore, that the Amazonian phaeopygus
group was not sampled here.
Another
paper (Batista et al. 2020) sampled ultraconserved elements (UCEs) across the
diversity of Turdus, including
multiple taxa in the assimilis/albicollis complex, but the previous
proposals noted that it, unfortunately, did not sample daguae. Batista et al. (2020) did sample UCEs from T. albicollis; however, both subspecies
included (phaeopygus and spodiolaemus) are part of the same
Amazonian subspecies group. UCE data indicated that T. albicollis and T.
assimilis are sister taxa but with a deep divergence. That UCE tree, from
their supplemental data, is shown below.

A
portion of supplemental Figure 2 from Batista et al. 2020, showing UCE-based
phylogenetic relationships in the albicollis/assimilis complex. The node separating albicollis from assimilis was estimated to be 3.16 Mya.
However,
the supplemental data from Batista et al. (2020) show that they did in fact
extract mitochondrial data from their UCE reads, and combined these with
existing mitochondrial DNA samples to estimate a phylogeny that does include
all four major clades in the complex. Although the standard mitochondrial gene
tree issues apply, they do point to deep divergences between all four groups
and resolve the taxon sampling issues raised previously.
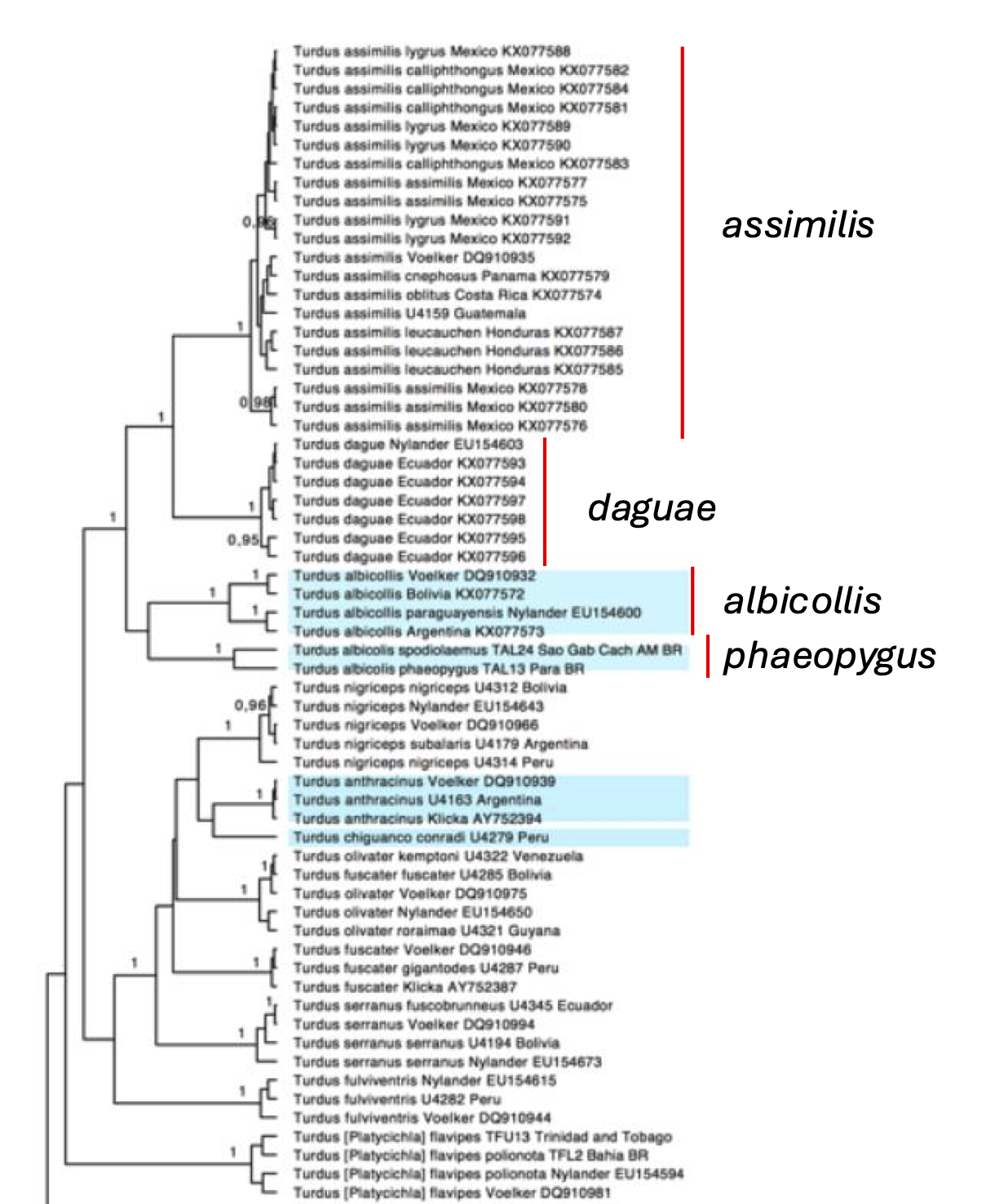
A
portion of Figure S5 from Batista et al. (2020) showing the phylogenetic
relationships in the Turdus assimilis/albicollis complex based on the mitochondrial gene cytb. We have highlighted the subspecies
groups of interest here with red bars. The scale bar on this mtDNA phylogeny
figure is difficult to interpret, as the scale bars below 5 Mya are missing,
but by our estimate, the node separating albicollis
from assimilis is approximately 4 Mya
(so, just older than from the UCE data) and the node separating daguae and assimilis is approximately 2.5 Mya. Of note, the node separating phaeopygus/spodiolaemus and nominate albicollis
is approximately 3.5 Mya. The samples highlighted in blue are those determined
by the authors to represent particularly deep intra-specific splits. Note that
the node ages of the four major clades in the assimilis/albicollis
complex are comparable to or older
than other well-established species in the genus.
Phenotypic variation
In
addition to the plumage differences noted in earlier proposals, Vallely and
Dyer (2018) mentioned that daguae
also show a dusky bill tip lacking in assimilis,
which they illustrated as having a solid yellow bill. However, online photos
show considerable variation in dusky coloration on the bill in assimilis, which may be age- or
sex-related. However, the few available photos of daguae show a considerably darker bill, solidly dark in almost all
cases. Herzog et al. (2016) illustrated phaeopygus
as having a solidly dark bill, versus a yellow bill with a dark tip in contemptus of the albicollis group. Photos online show phaeopygus having either a dark bill or a yellow mandible
contrasting with a dark maxilla.
Vallely
and Dyer (2018) illustrated both gray and brown birds for T. assimilis and noted that these populations are known from
adjacent localities in Honduras. Collar et al. (2024) illustrated the
subspecies T. assimilis atrotinctus
of the Caribbean slope of Honduras and Nicaragua as being dark gray and mention
in the text that the subspecies leucauchen
is also dark gray. In our search of online photos and field guide references,
it appears that the brown populations are found in most of Mexico and extend south
on the Pacific slope to southern Guatemala. These are nominate assimilis and some related subspecies,
which are paler and more uniformly brown than other subspecies. Dark gray birds
are found on the Caribbean slope from the humid slope of southeastern Mexico,
south through Guatemala, Honduras, and Nicaragua, and again in the mountains of
most of Costa Rica except the far south (nicely illustrated in Howell &
Webb 1995). In the southern Pacific slope of Costa Rica and through its range
in Panama, there is another brown subspecies, cnephosus, but this one is more contrasting and paler below than
the northern nominate brown subspecies. Most of the above phenotypic
differences were also noted by Hellmayr (1934), but we note that from the
mtDNA, these are all genetically very similar.
Lastly,
a “small and dark” subspecies coibensis
is known from Isla Coiba, but its genetic affinities are unknown. In his
description of coibensis, Eisenmann
(1950) gave the diagnosis as “closest to daguae”,
especially in the underpart and bill color, and that it is different than cnephosus in that regard. Wetmore (1957)
did say that coibensis differs from daguae in being “larger, more olive
above and grayer below, with the unmarked white area on the foreneck less in
extent”, so some plumage differences exist. Wetmore (1957) also provided
morphometrics for coibensis, which
could be compared with other taxa. Eisenmann (1950) used this plumage
similarity as evidence that daguae
was conspecific with assimilis, with coibensis as the geographic
intermediate. However, he also combined all these taxa under an expanded albicollis. Ridgely & Gwynne (1989) started
that coibensis have a blackish bill
and are ruddier above than cnephosus,
and also that it is the most numerous forest bird on the island, quite
different in this regard from the mainland populations. However, we now know
that Isla Coiba has some very interesting biogeography; for example, the
endemic Coiba Spinetail is most closely related to South American taxa, so we
think it is more likely that something interesting is going on with coibensis, possibly a future candidate
for species status. If daguae is
split, we think coibensis should be
tentatively retained with assimilis for
now, given that it is vocally much like the assimilis
group (see below). Some photos of this taxon are here: https://macaulaylibrary.org/asset/615988098 and here https://macaulaylibrary.org/asset/451105661
Regarding
morphometrics, Hellmayr (1934) noted that daguae
is notably smaller than other taxa. This was described in more detail in NACC
2022-A-4, but also illustrated nicely in the PCA plot from Núñez-Zapata et al.
(2016) shown below.
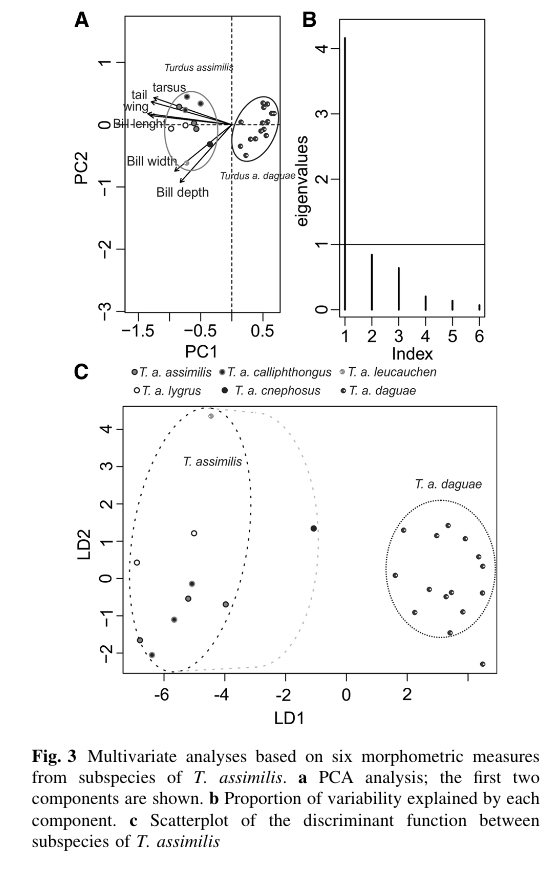
In
the T. albicollis complex, the main
phenotypic differences is a dark bill and gray flanks in members of the phaeopygus clade, and more yellow in the
bill and obvious rufous/cinnamon flanks in members of the albicollis clade. This pattern appears to hold even where the taxa
replace each other in Bolivia.
Vocal variation
|
|
|
|
|
|
|
|
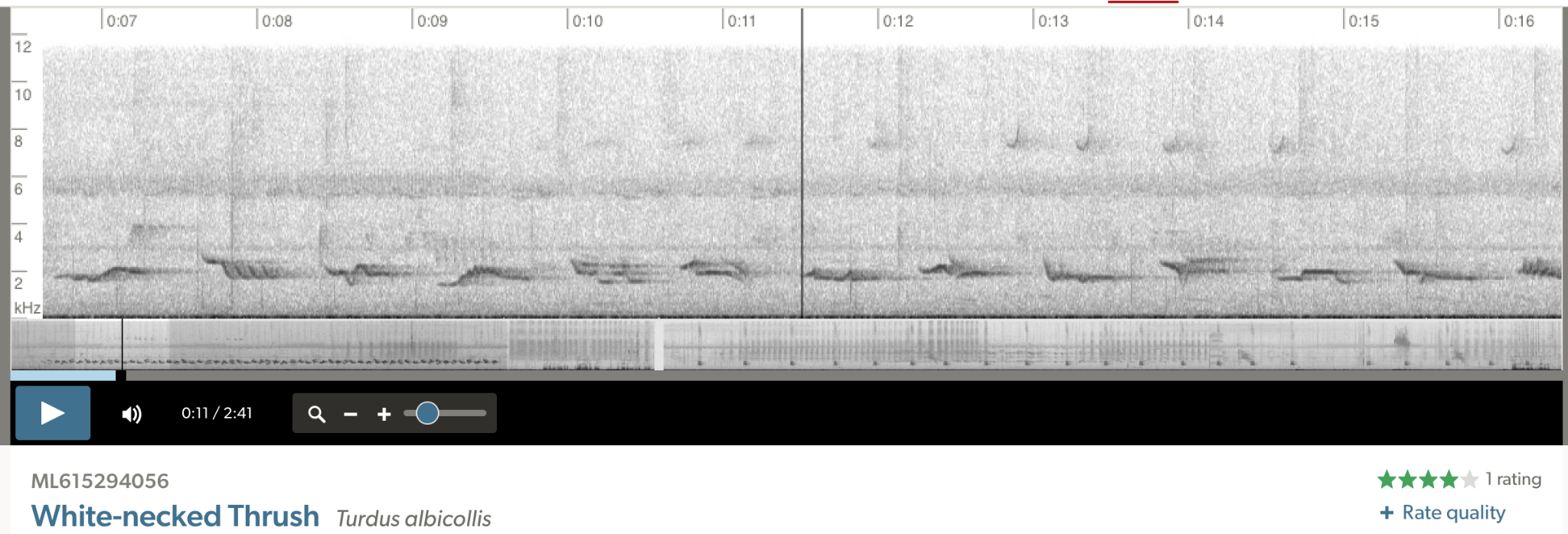



Comparisons
of song from the four major groups in the T.
assimilis / T. albicollis
complex. Top to bottom: 1) phaeopygus group,
2) nominate albicollis, 3) nominate assimilis, 4) daguae. Note the similar form and structure of daguae to T. albicollis.
Note that daguae seems to lack the
doubling of notes and the notes are more level.
To
our knowledge, the only quantitative analysis of vocal variation within the T. assimilis / T. albicollis / T. daguae complex comes from Boesman (2016), which
is worth reading: https://birdsoftheworld.org/bow/ornith-notes/JN100305
Below
are links to exemplar songs that highlight the differences mentioned by Boesman
(2016) and our comments on these differences. We note that Nacho Areta made
some excellent comments on the SACC proposal that support the distinct song of daguae. We agree that daguae sounds much more like Amazonian phaeopygus than like assimilis. Ridgely & Greenfield
(2001) had the following to say about the song of daguae: “Song a long-continued musical caroling with somewhat
monotonous effect similar to White-necked Thrush’s but pace a little faster
(very different from White-throated Thrush)”.
A
great example of most vocalizations in assimilis,
showing especially the distinctive song: https://macaulaylibrary.org/asset/72851
Here
is a good song from daguae: https://xeno-canto.org/275527 which certainly sounds
higher pitched than the rest to us and very different from assimilis. A few more examples here: https://xeno-canto.org/species/Turdus-daguae?view=3 and here:
Good
song example from phaeopygus group: https://macaulaylibrary.org/asset/158569851
Good
song example from nominate albicollis
group: https://macaulaylibrary.org/asset/615294056
The
one available song recording of coibensis
sounds typical of assimilis: https://macaulaylibrary.org/asset/69480131
Boesman
(2016) looked only at songs (which are, of course, critical), but there appear
also to be considerable differences in the calls. There are at least three main
call types in this clade: one whistled and longer, a short rough “churt” note,
and an odd chattering call.
The
whistled call note is clear and rising-falling in assimilis: https://macaulaylibrary.org/asset/231691361
But
has a rising emphasis at the end in daguae:
https://macaulaylibrary.org/asset/288967681
But
some assimilis may approach this: https://macaulaylibrary.org/asset/72851
Including
coibensis: https://macaulaylibrary.org/asset/69480121
This
call is much lower-pitched in Amazonian albicollis
(calls after 3:15 mark): https://macaulaylibrary.org/asset/188608
And
to be thorough, here is that call from nominate albicollis, which is also short and low-pitched: https://macaulaylibrary.org/asset/466693731
In
a cursory search, the short “churt” call sounds fairly similar across taxa, but
more work should be done here.
The
Amazonian taxa most commonly give the odd chattering repeated call, which is
uncommon or rare in other taxa. A good example from phaeopygus is here: https://macaulaylibrary.org/asset/245273
Here
is that call in assimilis: https://macaulaylibrary.org/asset/591450671, which sounds much
clearer and whistled than in phaeopygus,
mirroring the differences in the whistled call.
This
is the only example we can find for daguae:
https://xeno-canto.org/64330, which sounds very
different than assimilis and more
like phaeopygus, but with one
recording, it’s hard to be certain that this is a reliable difference.
The
call of coibensis is described as “a
guttural call like birds of the western highlands” and a “complaining chur-r-r or pru-rr-r” (Wetmore et al. 1984).
Distribution:
Turdus assimilis has a broad
elevational distribution, occurring in mid-elevations and low elevations from
northern Mexico through central Panama. This species is found in many foothill
localities in Costa Rica and Panama, being more widespread in the Pacific
lowlands than in the Atlantic lowlands. Farther south in this distribution, it
is found primarily at middle elevations but occasionally wanders to the
lowlands. In central Panama, it is found in the isolated hilly regions of Valle
de Antón, Altos de Campana, and Cerro Hoya (all west of the Canal zone). A few
eBird records from the Canal Zone and Cerro Azul represent the assimilis group based on photos. Ridgely
& Gwynne (1989) note that the birds in the Canal zone are wanderers from
elsewhere, with numbers peaking in November-January, and also mention that
“E.S. Morton found it to almost completely disappear from Cerro Campana during
the dry season”. So, it seems that small numbers of the foothill birds from
west of the Canal zone disperse eastward, including likely the Cerro Azul
records, and that Altos de Campana is the easternmost breeding population.
The
Pacific slope of the Darién is the northernmost extent of daguae. Wetmore et al. (1984) cite specimens from Cerros Pirre and
Tacarcuna, where they considered daguae
to be fairly common. They also mention a specimen from Cerro Sapo in the
coastal Serranía del Baudó. Ridgely & Gwynne (1989) also assign both the
Pirre and Tacarcuna birds to daguae.
In eBird, all records in the Darién south of the Chucunaque River appear to be daguae, including records in the
foothills of the Serranía de Pirre; records in the Cerro Tacarcuna lack photos.
Of interest are eBird records on the Cerro Chucantí in western Darién. Just as
we were wrapping up this proposal we noticed that a “Turdus assimilis” was marked as a background species in a recording
from this site, and it sounds to us like a typical daguae, thus extending the distribution of this taxon slightly
westward: https://xeno-canto.org/2974. Thus, it appears that
assimilis and daguae are spatially isolated by intervening lowlands in central
Panama (specifically, the lowlands around the Río Chepo),with assimilis extending as far south and
east as Cerro Azul, where they occur around 800 m, and daguae extending as far north and west as Cerro Chucantí, where
most records are > 700 m (with one record at 100 m without any
documentation). The highlands of central and southern Panama are connected by
the Serranía de San Blas, but we are not aware of records of either assimilis or daguae from this region; notably, this mountain range has a high
elevation of ~748 m, lower than the elevations of both assimilis and daguae on
the most adjacent mountains to the “gap” between these taxa and perhaps not
suitable for populations of either taxa. Thus, we find no evidence for sympatry
within the complex. .
Regarding
coibensis, Wetmore et al. (1984) cite
specimen records from some of the islands between Coiba and the mainland,
namely Isla Brincanco and Isla Rancheria, so it seems that this subspecies
approaches the mainland. However, this taxon is found down to sea level, even
in mangrove swamps, unlike foothill cnephosus
(Wetmore et al. 1984)
Possibly
of relevance, the eBird science map has different abundance patterns for the
two taxa, with daguae being uncommon
and assimilis being common. However,
there are few occurrence records for daguae,
which may impact the reliability of this difference. However, Ridgely &
Gwynne (1989) do state that daguae is
uncommon, while cnephosus is fairly
common.
In
the T. albicollis complex, most
members of the phaeopygus clade are
widely separated from those of the albicollis
clade by the cerrado of Brazil and Bolivia. However, contemptus of the albicollis
clade is found on the east slope of the Andes from southern Puno (Peru) through
most of Bolivia into extreme northern Argentina (eBird records, Birds of the
World, Herzog et al. 2016). In southern Peru and northern Bolivia, it is
parapatric (elevational replacement) with members of the lowland phaeopygus group. From the mitochondrial
phylogeny (Batista et al. 2020), the split between these two clades (albicollis and phaeopygus) is about 3.5 Mya.
Effect on SACC area:
Splitting
T. daguae from T. assimilis would result in no additional species for the SACC
area, as assimilis is extralimital.
Splitting T. phaeopygus from T. albicollis would result in one
additional species for the SACC area.
Recommendation:
Based
on differences in Cytb,
morphometrics, voice, and plumage, we posit that the T. assimilis/T. albicollis complex
as a whole comprises either one broad-ranging taxa with very
well-differentiated subspecies or four species-level taxa: T. assimilis in the north, T.
daguae in the Chocó, T. phaeopygus
in the Amazon, and T. albicollis in
southern South America. Given concordant differences in genetics, plumage, and
(in some cases) song for each of the groups, we recommend a YES vote on
elevating both daguae and phaeopygus to species rank.
Regarding
names, we suggest that the committee members read the previous NACC proposal
and comments on both the NACC and SACC proposals. Clements/eBird lists daguae as White-throated Thrush (Dagua),
and Dagua Thrush is used by Ridgely & Greenfield (2001). Hilty & Brown
(1986) also state that daguae has
been considered a separate species by others, under the name Dagua Thrush. So,
there is historical usage of this name. The name is based on the collecting
locality, the Rio Dagua, which is a fairly small river, but the name is
memorable and unique, and there is plenty of precedence for using the
collecting locality for the species name (e.g., Altamira Oriole, Tennessee
Warbler), although those names are often criticized for not being particularly
useful. Choco Thrush is a logical choice, given that it is endemic to this
biogeographic region. There are, however, plenty of other birds with the
“Choco” name, and two other Turdus
are endemic or near-endemic to the Chocó region (T. obsoletus and T.
maculirostris). However, of these three species, the range of daguae most closely matches that of the
bioregion. We therefore lean towards Dagua Thrush. This name should be
considered in consultation with NACC.
For
T. phaeopygus and T. albicollis, Clements/eBird uses
Gray-flanked for phaeopygus and
Rufous-flanked for albicollis, which
are acceptable and available names. Other options could include retaining
White-necked for albicollis and
adopting a new name for phaeopygus,
but it is not clear what other names might apply to that bird. Amazonian Thrush
is an option, as the species is widespread in the Amazon Basin, but it is one
of many Amazonian Turdus species. We
encourage SACC members to discuss potential names for these taxa.
Please
vote on the following:
A. Elevate daguae
to species rank. We recommend a YES
vote.
B. Elevate phaeopygus
to species rank. We recommend a YES
vote.
Literature Cited:
Batista, R., Olsson,
U., Andermann T., Aleixo A., Ribas C.C. and Antonelli, A. 2020. Phylogenomics
and biogeography of the world's thrushes (Aves, Turdus): new evidence for a more parsimonious evolutionary history.
Proc. R. Soc. B. 28720192400 http://doi.org/10.1098/rspb.2019.2400
Boesman, P. 2016. Notes
on the vocalizations of White-throated Thrush (Turdus albicollis). HBW Alive Ornithological Note 305. In: Handbook
of the Birds of the World Alive. Lynx Edicions, Barcelona. (retrieved from
https://birdsoftheworld.org/bow/ornith-notes/JN100305).
Collar, N., J. del
Hoyo, J. S. Marks, and G. M. Kirwan (2024). White-throated Thrush (Turdus assimilis), version 1.1. In Birds of
the World (S. M. Billerman, B. K. Keeney, P. G. Rodewald, and T. S.
Schulenberg, Editors). Cornell Lab of Ornithology, Ithaca, NY, USA.
https://doi.org/10.2173/bow.whtrob1.01.1
Eisenmann, E. 1950.
Some notes on Panamá birds collected by J. H. Batty. The Auk 67(3):364-367.
Hellmayr, C. E. 1934.
Catalogue of birds of the Americas, part XI. Field Museum of Natural History
Zoological Series Vol. XIII. Chicago, USA.
Herzog, S.K., Terrill,
S.T., Jahn, A.E., Remsen, Jr., J.V., Maillard Z., O., García-Solíz, V.H.,
MacLeod, R., Maccormick, A., and Vidoz, J.Q. 2016. Birds of Bolivia Field
Guide. Asocación Armonía, Santa Cruz de la Sierra, Bolivia.
Hilty, S.L., and Brown,
W.L. 1986. A guide to the birds of Colombia. Princeton University Press.
Howell, S.N.G., and
Webb, S.W. 1995. A guide to the birds of Mexico and northern Central America.
Oxford University Press.
Núñez-Zapata, J.,
Townsend Peterson, A. and Navarro-Sigüenza, A.G. 2016. Pleistocene
diversification and speciation of White-throated Thrush (Turdus assimilis; Aves: Turdidae). Journal of Ornithology
157:1073–1085. https://doi.org/10.1007/s10336-016-1350-6
Peters, J. L. 1964.
Check-list of birds of the world. Vol. X. (E. Mayr & R. A. Paynter, Eds.).
Museum of Comparative Zoology. Cambridge, Mass.
Ridgely, R.S., &
Greenfield, P. J. 2001. The birds of Ecuador. Cornell University Press.
Ridgely, R.S., &
Gwynne, Jr., J. A. 1989. A guide to the birds of Panama, with Costa Rica,
Nicaragua, and Honduras. Princeton University Press.
Vallely, A. C. and D.
Dyer. 2018. Birds of Central America. Princeton University Press.
Wetmore, A. 1957. The
birds of Isla Coiba, Panamá. Smithsonian Miscellaneous Collections, publication
4295.
Wetmore, A., Pasquier,
R. F., and Olson, S.L. 1984. The Birds of the Republic of Panamá. Part
4.-Passeriformes: Hirundinidae (Swallows) to Fringillidae (Finches).
Smithsonian Miscellaneous Collections, v. 150.
Oscar Johnson, and
Jacob C. Cooper, May 2025
Note on English names from Remsen: For daguae, if
the proposal passes I suggest we go ahead and use Dagua Thrush (see discussion
and rationale above). This is the name
already in use in several places, even going back to Hellmayr (1934) as the
name for the subspecies; thus, it is broadly acceptable, and so in the
interests of stability I see no point in trying to find a “better” name at this
point through a separate proposal. If
anyone objects to this, speak out. NACC
has already endorsed Dagua Thrush (although it is extralimital to NACC area.
For phaeopygus,
we will need a separate English name proposal, which could involve creating new
names for both daughter species. Note
the discussion above for potential names for phaeopygus and let those
incubate.
Voting Chart: https://www.museum.lsu.edu/~Remsen/SACCPropChart1044+.htm
Comments from
Bonaccorso:
“A. YES. It’s a shame there is no nuclear evidence
available, but the deep mtDNA divergence between T. daguae and T.
assimilis, along with what appear to be significant differences in song,
supports the case for treating them as separate species. I wouldn’t place too
much emphasis on the morphometric data alone, but when considered alongside the
mitochondrial genetic and vocal differentiation, it does provide stronger
support for the decision to separate these two taxa. The plumage differences
are subtle, but that is also true for the other species within the group.
“B. YES. In this case,
both mitochondrial data and UCEs (albeit with only one sample from each form)
support the separation of these two taxa. The songs also sound very different
to me—though I acknowledge that I am not an expert in bioacoustics.”
Comments from Stiles: “YES to A and B: I
consider that Elisa’s arguments sum up nicely the evidence.”
Comments from Naka:
“A) YES. I think the
evidence for splitting daguae based on phenotypical, vocal, and genetic
data are very compelling. I am happy with Dagua Thrush as the English name.
“B) YES. I like this
split, which is well supported by morphological and molecular data. I suspect
that new studies may suggest the need of further subdivisions in these two
clades, but I think it is an advancement to recognize the Amazonian and Eastern
forms as different species. As for the English names, I have nothing against Gray-flanked
for the phaeopygus group and Rufous-flanked for the albicollis group,
as suggested in the proposal.”
Comments from Rafael D.
Lima (guest voter):
“YES to A and B. While additional data are certainly needed to definitively
establish species limits within this complex, I believe the most appropriate
taxonomic treatment at present is to recognize these four groups as separate
species, given the pieces of evidence described in the proposal. It is worth
noting that the distributions of the albicollis and phaeopygus groups
may not be as allopatric as depicted in HBW/BOW or described in the proposal.
Numerous photographic records from central Brazil suggest potential contact in
that region (see https://www.wikiaves.com.br/mapaRegistros_sabia-coleira), mirroring patterns
seen in other Amazonian vs. Eastern taxa pairs (e.g., Camptostoma obsoletum
obsoletum/C. o. napaeum, Thraupis episcopus/T. sayaca,
Leistes militaris/L. superciliaris). Additionally, some authors
have proposed that T. a. contemptus may represent a hybrid population
between albicollis and phaeopygus, based on its intermediate
plumage (McCarty's 2006 Handbook of Avian Hybrids of the World, p. 238). This
seems plausible, and future sampling in all these putative contact zones will
be crucial for clarifying the species boundaries in this group.”
Comments
from Areta:
“A. Yes to T. daguae.
I was convinced of this split in the previous proposal for the reasons given in
my previous vote and continue to be persuaded after reading this proposal.
“B. I am hesitating on
this one. The deep split, plumage differences, and presumed parapatry
between contemptus (a rather grayish
or brownish-flanked western taxon of the albicollis
group) and spodiolaemus are
indicative of speciation. However, I am underwhelmed by the vocal differences,
the vocalizations are similar and clearly indicate that phaeopygus and spodiolaemus
are part of the same clade as albicollis,
at least based on their songs. What about calls? It also worries me what to do
with phaeopygoides. This taxon would
belong to the phaeopygus group based
on plumage (and geography?) but is missing vocal assessment (listen to
https://macaulaylibrary.org/asset/69474 and
https://macaulaylibrary.org/asset/177172 for example, clearly part of the albicollis/phaeopygus clade) and genetic
samples, and it would occur to the north ("northeastern Colombia to
northern Venezuela, and Trinidad and Tobago"), therefore widely separated
from the albicollis group. Because it
is largely montane, it would not fit comfortably with the "lowland
Amazonian" concept of phaeopygus
put forward in the proposal. Note also that phaeopygus
occurs across the Guianan region.”
Comments from Claramunt: “YES to both A and B. These are
clearly four well differentiated taxa. There seem to be no evidence of
reproductive compatibility or hybrids that suggest they a single species.”
Comments from Robbins: “YES, to both A & B, as all
data sets, genetic, plumage, and voice support recognition of daguae and
phaeopygus as species.”
Comments
from Remsen:
“A. YES. My previous
reservations on this split are now satisfied by additional vocal and genetic
data.
“B. NO. It is tempting
to vote yes on this one, for the reasons outlined in the proposal and also in
many of the comments, and I think the phaeopygus group will be shown
conclusively to be a good species eventually.
The data are indeed consistent with species rank for the most part,
although the plumage differences have no bearing on species rank because
different subspecies also have to have diagnostic plumage differences. The vocal data are weak and are screaming for
a forma analysis of songs and calls. The genetic results are based on a single
sample each of phaeopygus and spodiolaemus. As Nacho pointed out, phaeopygoides was not sampled vocally or
genetically. Andean contemptus also needs to be included in
these analyses; Nuñez et al. had samples, so why were these not also included
in Batista et al.? The presumed cases of parapatry could use a more through
analysis, especially because they would be indisputable evidence for species
rank if thoroughly documented. The
proposal did a great job of trying to patch the holes in the data to propose a
coherent taxonomy, but there are just too many holes, in my opinion, to make a
major change to the status quo. What’s
the rush? Let’s make sure we have it
right.”
Comments
from Zimmer:
“A) “YES. The case for splitting daguae from assimilis,
based upon genetic data (deep mtDNA divergence); song differences, which,
although not quantified in any comprehensive analysis, are qualitatively
obvious to the human ear, and visually distinct in spectrograms; and phenotypic
differences in plumage and morphometrics; are compelling.
“B) “YES. Van makes some good points regarding the weak
points in the data sets, and I agree with Nacho that the song differences
between phaeopygus and albicollis are underwhelming. However, the distinctive chattering call that
is common in phaeopygus is not something that I hear from Atlantic
Forest albicollis, which, in my experience, give the repeated short call
(e.g. https://macaulaylibrary.org/asset/466693731), both as spontaneous
calls, and, in response to playback.
That, combined with the mtDNA & UCE data supporting deep divergence
between all 4 clades (assimilis-group, daguae, phaeopygus-group,
and albicollis-group); obvious and consistent plumage and bare parts
differences; and the presumed parapatry between contemptus and spodiolaemus,
as well as the possible parapatry in central Brazil alluded to by Rafael Lima,
are enough to sway me. Obviously, more
data are needed, particularly from potential contact zones, to fully resolve
species-limits in the whole assimilis-albicollis complex, and I would
especially like to see the status of contemptus and phaeopygoides
clarified (as well as that of coibensis, even though it is outside of
our purview). Like Luciano, I suspect
that additional research will not only confirm the current proposed splits but
will also support further splits within the complex. But for now, I think the proposed splits
better reflect the relationships based on the evidence in front of us.”
Comments from Lane: “A) YES. B) YES. I understand the reservations of Van and Nacho, but
to me, the voices of the two taxa I know best (spodiolaemus and contemptus) are so different from one another, that they are clearly not
conspecific in my mind. More eastern albicollis is perhaps a bit closer to the Amazonian group’s voice, but the calls,
as Kevin mentions, still distinguish it from them easily. Overall, between
size, plumage, and voice, the southeastern T.
albicollis (including contemptus) and the Amazonian phaeopygus (including spodiolaemus and phaeopygoides) are plenty distinct and if daguae is to be recognized, I think the evidence available requires these to
be as well.”