Proposal (940) to South
American Classification Committee
Recognize Pseudocolaptes
johnsoni as a separate species from P. lawrencii
Pseudocolaptes johnsoni was
described as a new species by Lönnberg and Rendahl in 1922 based on a specimen
reportedly from Baeza, Ecuador (ca. 1800 m on the Eastern slopes of the Andes).
Subsequently, Zimmer (1936) found four specimens that matched perfectly
Lönnberg & Rendahl’s description of P. johnsoni but collected
on the Western slopes of the Andes. The match was so striking that Zimmer
(1936) not only concluded that johnsoni inhabits the western slopes but
also cast doubts about the provenance of the type specimen, as all other
specimens reported from that region are clearly boissonneautii. Zimmer
(1936) also made clear that the distinctive plumage of johnsoni cannot
be confused with the juvenal plumage of boissonneautii. Zimmer ended up
considering johnsoni as a subspecies of P. lawrencii
instead, but without providing any evidence other than presumed “closer
affinities”.
Based
on plumage differences and elevational preferences Robbins & Ridgely (1990)
suggested that johnsoni deserves species status, a treatment followed by
few lists (Ridgely & Tudor 1994, Ridgely & Greenfield 2001, del Hoyo
& Collar 2016). A previous SACC proposal based on this evidence was
rejected because of lack of additional evidence such as vocalizations: https://www.museum.lsu.edu/~Remsen/SACCprop28.htm
A
more recent proposal was based on results of playback experiments showing that
the song of johnsoni does not elicit any response in lawrencii
individuals, suggesting significant vocal differences potentially producing
premating isolation (Freeman & Montgomery 2017):
https://www.museum.lsu.edu/~Remsen/SACCprop754.htm
The
analyses of Spencer (2011) and Boesman (2016) show the differences in song
characteristics between johnsoni and lawrencii. If the samples
are representative of each taxon, they show well-marked differences:
“johnsoni: Song is a high-pitched rattled series
of notes slowing into stuttering and ending (always) with a characteristic
high-pitched down-slurred note.”
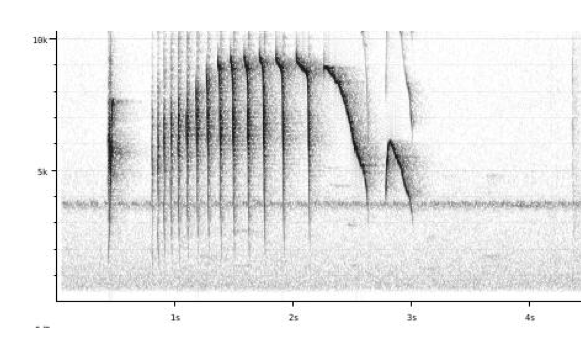
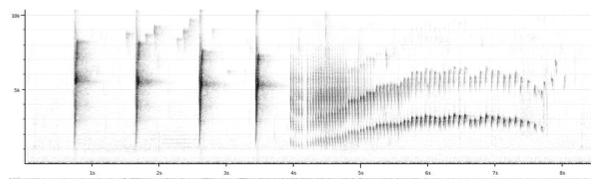
“lawrencii:
Song is a number of well-spaced staccato notes (always present unlike johnsoni)
followed by a trill, which usually first ascends in pitch and then slightly
descends while slowing down in pace.” (Boesman 2016)
New
Information:
Forcina
et al. (2021) revisited the issue of the species status of lawrencii by
reanalyzing DNA sequences from a previous study (Derryberry et al. 2011). They
claimed that researchers so far have “neglected” the DNA evidence that
indicates that johnsoni is a separate species. They found that the single
sample of johnsoni showed levels of divergence comparable to those
between lawrencii and boissonneautii. In other words, the three
lineages diverged nearly simultaneously about 2 million years ago (using
mitochondrial clocks; compare to ~ 4 Ma in Derryberry et al. 2011). The
analysis could not resolve whether johnsoni is closer to lawrencii
or to boissonneautii. They discuss all the evidence accumulated so far,
including morphology, vocalizations, and habitat, and conclude that johnsoni
should be elevated to species status.
Maximum
likelihood analysis (Forcina et al. 2021 fig 1)
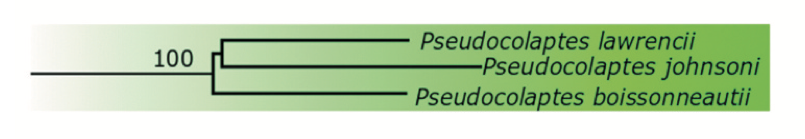
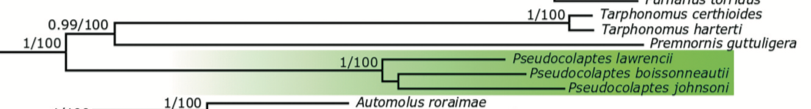
Bayesian
analysis (Forcina et al. 2021 fig 1)
Discussion:
The
data needed for evaluating the species status of johnsoni has
accumulated slowly over decades, a little at a time, mostly due to the rarity
of the species. But all pieces of evidence suggest that johnsoni is a
distinctive species. It is clearly diagnosable by plumage; it is not a cryptic
species by any means. I don’t understand why Forcina et al. claim that this is
a case of “cryptic diversity.”
Photographs
of the type specimen in the Stockholm museum are now publicly available here:
And
the photographs confirm the original description and, in my opinion, the match
between the type specimen and the AMNH series of 4 birds from W Ecuador.
Photographs
available online of live birds may be a bit confusing because of the artificial
variation in color produced by different light conditions and digital
adjustments, and the presence of birds in juvenal plumage, but here are what I
consider good representatives of adult johnsoni (very similar the AMNH
specimens):
https://macaulaylibrary.org/asset/178142751?_gl=1*1pz6d24*_ga*NjE4NTUxOTA5LjE2NDQ5NzU5MDk.*_ga_QR4NVXZ8BM*MTY0NzQwNzI3NC4yNS4xLjE2NDc0MDczMjkuNQ..#_ga=2.131747151.1345091137.1647401651-618551909.1644975909
https://macaulaylibrary.org/asset/41029281?_gl=1*30cct8*_ga*NjE4NTUxOTA5LjE2NDQ5NzU5MDk.*_ga_QR4NVXZ8BM*MTY0NzQ1Mjg5NS4yNi4xLjE2NDc0NTMwMTQuNDQ.#_ga=2.201353681.1345091137.1647401651-618551909.1644975909
In
both the holotype and the photos, note the deep rufous tones on back and flanks
(versus brown in the other two taxa), no light streaks on the mantle (vs. streaked
in boissonneautii), whitish “cheeks” (vs. buffy in lawrencii),
dark lower throat (not forming a gular band that is continuous with the light
cheeks like in the other two taxa), inconspicuous superciliary stripe (versus
thin but well-demarcated stripe in the other two taxa), blackish breast with
white rhomboid spots and rest of the belly rufous (versus predominating buffy
spots that coalesce to form a light-colored central belly in the other two
taxa).
The
song is clearly distinctive and not recognized by lawrencii in playback
experiments (Freeman & Montgomery 2017). For both plumage and
vocalizations, a more detailed analysis of geographic variation across the
Colombian Andes would have been desirable but I think that the evidence is
compelling.
Finally,
the genetic evidence adds a bit of additional information. But contrary to what
the title in Forcina et al. suggests, it doesn’t “untangle” anything. Genetic
data for just three individuals do not tell much about species limits. A random
sample of three individuals from a single large and old population can produce
a tree similar to the one found by Forcina et al. The species delimitation
algorithm based on the Poisson Tree Processes (PTP) used by Forcina et al. is
basically a mathematical formalization of a genetic divergence
threshold/yardstick criterion, in itself a rather weak species delimitation
criterion. The method classifies the nodes of the tree into speciation events
and coalescent (intraspecific) events based on levels of divergence. It thus
formalizes the observation that the three Pseudocolaptes are rather
divergent.
The
recovered topology doesn’t help much either. A strongly supported sister
relationship between johnsoni and boissonneautii would have
clearly made the case for separating johnsoni from lawrencii, but
there were conflicting topologies across methods, and clade support was nil. In
sum, the genetic data is consistent with the species status of johnsoni,
but the evidence is rather weak.
In
any case, I think that plumage, songs, and genes together provide sufficient
evidence suggesting that johnsoni deserves species-level status and
there is not a hint of evidence that suggest that johnsoni is conspecific
with lawrencii or with boissonneautii.
Recommendation:
I
recommend the treatment of johnsoni as a separate species.
Boesman, P. 2016. Notes on the
vocalizations of Buffy Tuftedcheek (Pseudocolaptes lawrencii). HBW Alive
Ornithological Note 87. In: Handbook of the Birds of the World Alive. Lynx Edicions, Barcelona.
Del Hoyo, J., & N. J. Collar 2016. Illustrated
checklist of the birds of the world, Volume 2: Passerines. Lynx Edicions,
Barcelona, Spain, & BirdLife International, Cambridge, UK.
Forcina, G., P.
Boesman, & M. J. Jowers 2021. Cryptic diversity in a Neotropical avian
species complex untangled by neglected genetic evidence. Studies on Neotropical
Fauna and Environment: 1-8. https://doi.org/10.1080/01650521.2021.1915674
Freeman, B. G., G. A.
Montgomery. 2017. Using song playback experiments to measure species
recognition between geographically isolated populations: a comparison with
acoustic trait analyses. Auk. 134(4):857–870.
Lönnberg, E., & H.
Rendahl 1922 A contribution to the ornithology of Ecuador. Arkiv för Zoologi
14(25): 1-87.
Ridgely, R. S. & P.
J. Greenfield. 2001. The birds of Ecuador. Vol. I. Status, distribution, and
taxonomy. Cornell University Press, Ithaca, New York.
Ridgely, R. S. & G.
Tudor. 1994. The bird of South America. Vol. II. The suboscine passerines.
University of Texas Press, Austin, Texas.
Robbins, M. B., &
Ridgely, R. S. 1990. The avifauna of an upper tropical cloud forest in
southwestern Ecuador. Proc. Acad. Nat. Sci. Philadelphia 142: 59-71.
Spencer, A. 2011.
Variation in tuftedcheek vocalizations. https://www.xeno-canto.org/article/99
Zimmer, J. 1936.
Studies of Peruvian birds, no. 21. Notes on the genera Pseudocolaptes, Hyloctistes,
Hylocryptus, Thripadectes, and Xenops. American Museum
Novitates 862: 1-25.
Santiago Claramunt,
March 2022
Note from Remsen on English names:
After internal discussion with SACC members, it seems best to stick with Pacific
Tuftedcheek, despite the marine connotation, because this is the name used at
the onset of the suggestions to treat this as a separate species, and because
several other bird species in the region are also referred to as Pacific
Something, with the understanding that this refers to the Pacific side of the
Andes. Therefore, we think that no
formal proposal is needed.
_______________________________________________________________________________________
Comments
from Robbins:
“YES for recognizing Pseudocolaptes johnsoni
as a species. As we pointed out in our
1990 paper, we suspected that johnsoni deserved species status based on
plumage and elevational differences. In
the original SACC proposal, I voted against recognizing it as a species because
of the lack of vocal data. However,
given that Spencer (2011) and Boesman (2016) have established vocal
differences, I now vote to elevate it to species status. As pointed out by Santiago, I consider the
genetic data equivocal.”
Comments from Stiles: “YES. As noted by Santiago,
genetics, distributions, plumage, and vocalizations all support recognition of johnsoni
as a species (the genetic evidence is weakest, but nothing suggests that johnsoni
is NOT a distinct species).”
Comments from Areta: “YES. Vocal and plumage data
support the split. The playback experiments also support this, even when these
lack rigor. Genetic data is conflicting, but show a relative deep divergence
between lawrencii, boissonneautii and johnsoni. Harvey et al. (2020) recovered a sister relationship
(with low support) of boissonneautii
and johnsoni with lawrencii sister to them:”

Comments from Lane: “YES. Between the distinctive
vocalizations and the likelihood that P. lawrencii and P. johnsoni
are not sisters, it seems the only reasonable move to separate the two at the
species level.”
Comments from Pacheco: “YES. Accumulated evidence
including genetics, morphology, vocalizations, and habitat support the
recommendation of this proposal.”
Comments from Bonaccorso: “YES. All the evidence points
towards species status. There are distinctive plumage differences between P.
lawrencii, P. johnsoni, and P. boissonneautii (still, this
level of plumage differences alone does not grant more than subspecies status
in other groups). The difference in song is clear, and the genetic
differentiation (although from just one specimen per species) is deep. Even if
we cannot know the sequence of splits between all three species, which seem to
have happened very fast in evolutionary time, the available evidence points to
three lineages that have been evolving independently from each other for about
2-4 million years. Also, I know that allopatry is frequently used against
granting species status, but I love seeing a clear geographic break between P.
lawrencii and P. johnsoni.”
Comments from Remsen: “YES, barely. I think burden-of-proof is now on treating
them all as conspecific, but not entirely because of the reasons in the
proposal. Although all the relatively
new evidence points towards recognition of johnsoni as a species, my
rationale derives from the phylogenetic hypothesis in Harvey et al. (2020), as
shown in Nacho’s comments above.
Assuming that additional taxon-sampling would not affect the topology
(see further comments below) and assuming that the weakly supported sister
relationship of johnsoni and boissonneautii has a fair chance of being correct, then treatment of johnsoni as a subspecies of lawrencii would be incorrect,
and treatment of it as a subspecies of boissonneautii is also incorrect because these two taxa are close to being
elevationally parapatric in the Western Andes without any physical barriers
between them --- those two have to be treated as separate species. Add the fragmentary vocal data to this, and
in my opinion, johnsoni has to be
treated as a species until shown otherwise.
“However, the negative wording and implications in the Forcina et
al. paper merit a critical response here lest those who don’t know the details
take them at face value. First, the
title of the paper is an in-your-face, unjustified swipe at Derryberry et al.
(2011). The title proclaims that we ‘neglected genetic
evidence’, and that Forcina et al. have come to the rescue to “untangle it”. Their abstract also proclaims:
‘We
assemble already available mitochondrial and nuclear DNA sequences to assess
their taxonomy and to provide appointed committees with specific proof [italics
inserted by me] to ascertain the number of Pseudocolaptes species.’
“Where to start?
First, Derryberry et al.’s paper was a conceptual paper in which any
taxonomic comments would have been inappropriate and edited out by reviewers,
and so this paper is not to blame for “neglecting” the issue of johnsoni. Even so, there would have been good reasons
to “neglect” the genetic data. They were
based on an analysis of 6 neutral loci of only 3 individuals, including only
1 of the 8 subspecies of P. boissonneautii. Further, the unsampled
nominate subspecies would have to be included in any analysis to anchor any
taxonomic conclusion; nominate boissonneautii is also the taxon geographically closest to the range of johnsoni, and thus its absence is an additional reason to be
cautious about the interpretation of the existing results. It would have been just as unwise for SACC or
NACC to have reached any resolution on species rank for johnsoni on such weak data
as it would have been for Derryberry et al. to have commented on it. No amount of ex-post-facto reanalysis by
Forcina et al. of DNA sequences from those three samples (the only new data
presented in Forcina et al.) will fix that crippling problem. Rather than “untangling” the “neglected”
data, their oversimplification of the problem is a potential setback. As for “proof”, the only thing they “proved”
is that they don’t understand the problems of using genetic data in a complex
system, e.g. highly polytypic P. boissonneautii is not a panmictic gene pool or that failure to include the nominate
subspecies in an analysis prevents any firm taxonomic conclusions.
“Forcina et al.’s Abstract stated:
‘However, even when multiple lines of
evidence lean toward lumping or splitting of species, some taxonomic committees
refuse to acknowledge their validity until convincing genetic evidence is produced
and integrated with other sources of data.’
“This attack on SACC and NACC requires a direct response. Contrary to Forcina et al.’s (2021)
hyperbolic claim to have restored sanity to taxonomy with resounding evidence, the
data are borderline. The plumage
differences, for example, tell us only that they are diagnosable taxa, not
necessarily species under the BSC. Many
subspecies that are more distinctive in plumage than these three taxa intergrade
at contact zones and thus show that plumage differences themselves are seldom
isolating mechanism. As for the
vocalizations, the N is small, the playbacks are not a controlled experiment,
and variation within boissonneautii and lawrencii
was noted by Spencer (2011) as adding some complexity to interpretations of
inter-taxon vocal differences:
“With such a small sample
size it's not possible for me to tell if this is a systematic difference or
just individual variation, but with what I can see it appears that populations
of Streaked Tuftedcheek vary as much vocally as Buffy do. The break appears to
be somewhere in Colombia, as at least some of the recordings from Colombia are
of the lower pitched song types, and songs I have heard in Ecuador and Peru
have also been of this type.”
“As for “refuse to acknowledge”, this reflects a misunderstanding
of the process by which those committees operate. To make a change in the classification
requires a proposal, which then has to be approved by a 2/3 majority. SACC rejected an earlier proposal before the
online summaries of vocal information by Spencer and Boesman. The “johnsoni problem” has been on our
publicly available list of proposals to be done for at
least a decade. As noted above, the
genetic data of Derryberry et al. and Forcina et al. is far from “proof” of
species rank for johnsoni, yet I pressured Santiago to write up a SACC
proposal. Does this constitute being
labeled as “refuse to acknowledge”?
“Forcina
et al. also stated:
“Derryberry
and coauthors did produce genetic data from an individual of the contentious P.
johnsoni but did not use such information in their phylogeny nor for any
comparative analyses. In spite of the overt differences in phenotypes, the SACC
(South American Classification Committee) along with the NACC (North American
Classification and Nomenclature Committee) of the AOS (American Ornithological
Society) and, as a consequence, the eBird and Clements checklists still treat P.
johnsoni and P. lawrencii as conspecifics.”
“First,
the taxon johnsoni has been treated as a subspecies of lawrencii for a century or so, including by Dickinson (2003), which is clearly
indicated as the starting point for the first SACC classification. And as Forcina et al.
noted later in their paper, a previous SACC proposal in 2003 rejected treatment
of johnsoni as a species because it was largely based
on anecdotal information. Second, “overt
differences in phenotype” are not sufficient grounds for treatment as a
separate species under any BSC-based classification, so the relevance of that
point is uncertain. Further, although
lacking modern analyses (see caveats in Remsen 2003 HBW Furnariidae account),
the 8 subspecies of P. boissonneautii also purportedly
have “overt phenotypic differences” among them --- otherwise, they would not be
treated as subspecies.
“Forcina
et al. then stated:
‘In
this study, we assemble available genetic data as a completion of multiple
lines of evidence to provide taxonomic committees with all the tools needed for
making a sound decision about the number and identity of Pseudocolaptes species,
thus eliminating the confusion arising from conflicting taxonomic treatments
and assisting future conservation actions.’
“This
over-confidence is unwarranted. The data
are still weak, with gaping holes and small N.
My vote on this is a tenuous YES only because continued inclusion of johnsoni
as a subspecies of lawrencii cannot be justified by the data in Harvey
et al. (2020), which was itself “neglected” by Forcina
et al. (published online in July 2021).
Also, I have confidence in the experience of Spencer and Boseman in
evaluating species-level vocal differences, although I worry that until all 8
subspecies of boissonneautii are included in vocal comparisons, we may
be premature in assessing vocal variation as well.
“Finally, Forcina et al. put SACC, NACC, and
Derryberry et al. on the defensive because their non-recognition of johnsoni as a species is implied
to be an obstacle to conservation.
However, the Chocó region has been recognized as a region of high
endemism for over a century (since Frank Chapman’s monographs) and as a
biodiversity hotspot since the term was invented decades ago. Whether johnsoni is recognized as a species does not affect those conclusions, much
less act as some pivot point, and any conservation measures that affect this
region will also affect johnsoni. Finally, in a seminal analysis
of Neotropical regions of endemism (Cracraft 1985 paper in the “Eisenmann” AOU Monograph volume), subspecies as well as
species were used to define those regions of endemism, as was done also in
Chapman’s monographs; thus, the taxonomic rank does not prevent johnsoni from being included
in analyses of endemism, nor does the current SACC-NACC treatment prevent
conservation measures from treating johnsoni as a species given that BirdLife International has treated it as a
species since 2016.”
Comments
from Jaramillo:
“YES – the vocal data help here.”