Proposal (969) to South
American Classification Committee
Note from Remsen: This proposal is circulating concurrently at NACC; I
have made minor editorial changes in this version.
Revise the taxonomy of Amaurospiza seedeaters:
(a) treat Amaurospiza
relicta as a
separate species from A. concolor,
and (b) treat A. concolor and A. carrizalensis as conspecific
with A. moesta,
or (c) treat A. aequatorialis as a separate species from A.
concolor,
or (d) lump the five taxa as subspecies of A. moesta
Description of the
problem:
This
proposal seeks to revise the taxonomy of Amaurospiza
seedeaters and contribute to the efforts of the Working Group on Avian
Checklists (WGAC) in reconciling global checklists. The genus Amaurospiza, as currently recognized, is
comprised of five taxa of blue seedeaters that show minor differences in
plumage coloration and body measurements. Recognition of the five taxa as
species or subspecies, including the species to which a subspecies belongs, varies
among global avian checklists (Table 1). Howard & Moore and eBird/Clements
coincide in the three species they recognize (concolor, carrizalensis, moesta), HBW-BL recognizes two species (relicta, moesta), and IOC recognizes four species (concolor, aequatorialis, carrizalensis, moesta). Classification by the NACC and the SACC agrees with Howard
& Moore and eBird/Clements.
Table 1. Current taxonomy of Amaurospiza seedeaters in four global
avian checklists. Classification by the NACC and the SACC agrees with Howard
& Moore and eBird/Clements.
|
Taxa |
Howard & Moore + eBird/Clements |
HBW-BL |
IOC |
|
relicta (Griscom, 1934) |
A. concolor relicta |
A. relicta |
A. concolor relicta |
|
concolor Cabanis, 1861 |
A. concolor concolor |
A. moesta concolor |
A. concolor concolor |
|
aequatorialis Sharpe, 1888 |
A. concolor
aequatorialis |
A. moesta
aequatorialis |
A. aequatorialis |
|
carrizalensis Lentino &
Restall, 2003 |
A. carrizalensis |
A. moesta
carrizalensis |
A. carrizalensis |
|
moesta (Hartlaub, 1853) |
A. moesta |
A. moesta moesta |
A. moesta |
The
five taxa in the genus Amaurospiza
are allopatric and distributed from central Mexico to northeastern Argentina
(Table 2). Two of the three subspecies within A. concolor (relicta and concolor) occur in the area covered by
the NACC, from central Mexico to Panama. The third subspecies of A. concolor (aequatorialis), and the species A.
carrizalensis and A. moesta, are found in South America, and,
therefore, are under the jurisdiction of the SACC.
Table 2. Geographic distribution of Amaurospiza seedeaters.
|
Taxa |
Distribution |
NACC |
SACC |
|
relicta |
Mts. of s Mexico
(s Jalisco to Guerrero, Morelos and Oaxaca) |
A. concolor Blue
Seedeater |
|
|
concolor |
Mts. of s
Mexico (Chiapas) to Nicaragua, Costa Rica and Panama |
A. concolor Blue
Seedeater |
|
|
aequatorialis |
Mountains of
SW Colombia (Nariño) to n Peru (Cajamarca) |
|
A. concolor Blue
Seedeater |
|
carrizalensis |
N Venezuela
(lower Río Caroni in Bolívar) |
|
A. carrizalensis Carrizal
Seedeater |
|
moesta |
Locally from
se Paraguay to e Brazil and ne Argentina (Misiones) |
|
A. moesta Blackish-blue
Seedeater |
Background:
Amaurospiza seedeaters are
Neotropical resident species generally associated with bamboo thickets and
dense understory (Lopes et al. 2011). They feed on arthropods and bamboo seeds,
flowers, petioles, and buds (Areta et al. 2023); in Costa Rica they prefer
greener and healthier leaves (Figure 1, Pablo-Castillo 2018).

Figure 1. Male of Amaurospiza concolor feeding on bamboo leaves in Costa Rica
(Pablo-Castillo 2018).
The
taxa within the genus Amaurospiza
have a convoluted history. Here is a brief summary of the five Amaurospiza taxa:
concolor
The
genus Amaurospiza was described along
with the species A. concolor based on specimens from Costa
Rica (Cabanis 1861). Ridgway (1901) measured one Panamanian specimen from the
Salvin-Godman collection; he noted that the species was found in both Costa
Rica and Panama.
Griscom
(1934) recognized three subspecies within concolor:
concolor from northwestern Costa Rica
(Miravalles and Tenorio); and two additional subspecies that he proposed: grandior from the humid Caribbean forest
of eastern Nicaragua, and australis from
southwestern Costa Rica to western Panama. Griscom provided measurements of relicta (placed in Amaurospizopsis) and the three taxa within concolor (grandior, concolor, australis). He found overlap among most of the measurements with
the exception of the wing of Amaurospizopsis,
which was larger than in any of the concolor
taxa (Table 3).
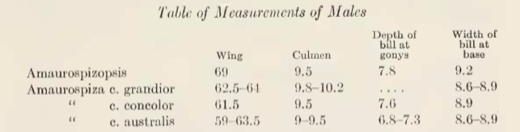
Table 3. From Griscom (1934), morphometric
measurements of male Amaurospizopsis
and Amaurospiza seedeaters.
Hellmayr
(1938) noted that grandior was
indistinguishable in color from concolor,
and that only one of three specimens had a slightly longer bill, suggesting
that grandior was not maintainable as
a separate taxon. The australis group
has not been reassessed since the original description by Griscom (1934), from
which it appeared that the main difference between australis and concolor
was the plumage coloration of the immature male (Figure 2). The two taxa grandior and australis are not recognized by any of the four global checklists;
they are currently grouped within concolor
concolor (Ramos-Ordóñez et al. 2020).

Figure 2. Original description of Amaurospiza concolor australis by
Griscom (1934).
moesta
The
taxon moesta was described as Sporophila moesta with a type from
Brazil (Hartlaub, 1853). Orr and Ray (1945) noted that Hellmayr (1904) found S. moesta to be identical to Amaurospiza axillaris (Sharpe 1888).
From then on, axillaris and moesta were synonymized and have been
considered part of the genus Amaurospiza.
Sharpe, in a key to the Amaurospiza
species, distinguished this taxon by its white axillaries, underwing coverts,
and quill-lining (Figure 3) [see below, that Hellmayr´s assessment indicates that
the immature male type of aequatorialis
lacks white underwing coverts as indicated by Sharpe, but that an adult male
has white underwing coverts, setting aequatorialis
apart from concolor; also see Table 4 in Areta et al. 2023]. However,
Sharpe also noted that taxa within Amaurospiza
are very closely allied, making it impossible to distinguish them from
descriptions alone. The taxon moesta
is mainly found in the Atlantic Forest, although records in pre-Amazonian
wooded habitats are recently increasing (Rising et al. 2020).

![]()
Figure 3. Key to Amaurospiza species from Sharpe (1888).
aequatorialis
Sharpe
(1888) described A. aequatorialis, a
species from the western foothills of the Andes of Ecuador, as a separate
species from concolor and moesta (under the name A. axillaris). This taxon is similar to concolor, but the forehead and eyebrows
are slightly paler bluish gray, and the bill is smaller. Hellmayr (1938)
treated aequatorialis as a subspecies
within concolor after examination of
four specimens, two from Ecuador and two from Colombia, and noted that aequatorialis was slightly smaller and
paler than concolor (Figure 4).

Figure 4. Note about Amaurospiza concolor aequatorialis by Hellmayr (1938).
AOU/AOS
considered aequatorialis as part of
the group A. c. concolor (AOU 1983;
AOU 1998). Although aequatorialis is
not mentioned explicitly in the checklist, the geographic distribution of A. c. concolor encompasses southwestern
Colombia and northwestern Ecuador. Recently, the taxon aequatorialis has also been recorded from northwestern Peru (Angulo
Pratolongo et al. 2012; Sánchez et al. 2012).
relicta
Griscom
(1934) described a new species within a new genus, Amaurospizopsis relictus, based on a specimen from Chilpancingo,
Guerrero, Mexico.
Hellmayr
(1938) suggested that relictus could
be a northern subspecies of concolor,
very similar in coloration, but with relictus
slightly larger with a deeper, stubbier bill.
Orr
and Ray (1945) compared Amaurospizopsis
and Amaurospiza, concluded that the
differences were not sufficient to warrant separate genera, and proposed that Amaurospizopsis be considered a synonym
of Amaurospiza. Griscom, in a letter
from 1944, concurred with Orr and Ray, mentioning that his views on avian
genera had changed since he proposed the genus Amaurospizopsis.
Orr
and Ray (1945) tentatively considered relictus
as a separate species, mainly due to the geographic hiatus between relictus and concolor, and the absence of intergradation in the specimens they
examined. They noted that in the length of the wing and the tail there is no
overlap between A. relictus and A. concolor, although the length of the
bill is the same for both species (Table 4). They reported that the color in
the adult male of relictus is grayer
and duller than the adult males of concolor.
Table 4. From Orr and Ray (1945), morphometric
measurements of Amaurospiza seedeaters.
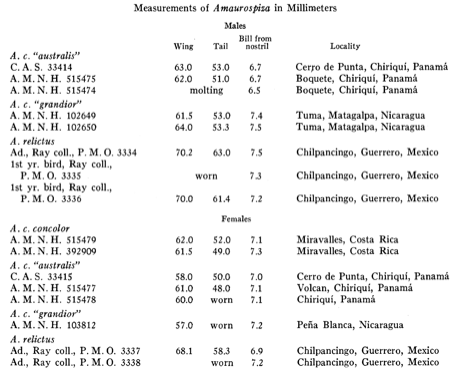
Miller
et al. (1957) noted under A. concolor
relicta: “Measurements of relicta … essentially bridge the size
gap between this form and A. c. concolor
of Central America and the color differences appear to be of a magnitude
frequent in races.”
AOU/AOS
considers relicta as a group within A. concolor (AOU 1983; AOU 1998). The
sixth edition (AOU 1983) noted: “The two groups are sometimes
recognized as distinct species, A.
relicta (Griscom, 1934) [Slate-blue Seedeater] and A. concolor [Blue Seedeater]”. The seventh edition (AOU 1998) mentioned the
two groups without referring to the possible recognition of two distinct
species.
Some
authors treat relicta as a separate
species (Eisenmann 1955; Davis 1972; Howell and Webb 1995). The song of relicta is described as similar to concolor but slightly higher and faster
(Howell and Webb 1995). Lentino and Restall (2003), considering bill shape,
size, color, and song differences, suspected that relicta might represent a separate species from concolor.
HBW-BL
split A. relicta from A. concolor based on the following
rationale:
“[relicta]
commonly treated as conspecific with A.
concolor; differs (in this analysis rictal bristles and nostrils accorded
equivalence of plumage characters) by its slate-blue vs. dark blue plumage in
male (2); longer rictal bristles (Griscom 1934) (allow 1); operculate nostrils
(Griscom 1934) (allow 1); shorter, deeper bill (allow 1); longer wing and tail
(mean of 3 male tails 57.7 mm vs mean of 5 males 50.8; allow 2); “slightly
higher and faster” song (Howell and Webb 1995) (at least 1).”
carrizalensis
The
taxon carrizalensis was described by
Lentino and Restall (2003) based on specimens collected on the river island
Carrizal in eastern Venezuela. The authors measured their specimen series and
specimens from the other taxa within the genus (Table 5). They found that carrizalensis has the longest bill and
most pointed wing of all the taxa within Amaurospiza.
Lentino and Restall diagnosed carrizalensis
as “separable from other members of the genus by the density of coloration and
black flammulations on the breast, overall size, wing formula, volume and shape
of the bill, and general measurements”. Lentino and Restall suggested that carrizalensis should be considered a
separate species, which was accepted by the SACC due to the large range
disjunction and morphological differences from concolor and moesta
(Proposal 74, https://www.museum.lsu.edu/~Remsen/SACCprop74.htm). Subsequently, the
English name Carrizal Seedeater was adopted by the SACC (Proposal 92, https://www.museum.lsu.edu/~Remsen/SACCprop92.htm).
Table 5. From Lentino and Restall (2003),
morphometric measurements of Amaurospiza seedeaters.
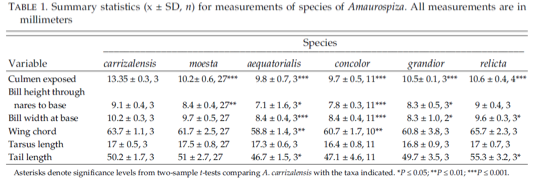
Additional
notes that involve more than one of the five taxa
Hellmayr
(1938) recognized three species: Amaurospizopsis
relictus, Amaurospiza concolor
(including grandior, concolor, aequatorialis), and Amaurospiza
moesta.
Orr
and Ray (1945) proposed that two races of Amaurospiza
concolor should be recognized: concolor
from Central America and aequatorialis
from northern South America. They added that further collecting efforts may
show relictus as a large, pale,
northern race of concolor.
Monroe
(1968) noted that, in Honduras, concolor
is a rare resident of the Caribbean lowlands, where it inhabits open rain
forests, forest edges, and second growth. Monroe examined a series of relicta and concluded that the Honduran
exemplars of concolor are not
conspecific with Mexican relicta.
Honduran concolor and relicta differ in morphology and
habitat, given that relicta inhabits
mountain ranges. However, the currently known elevational range of concolor in Central America is 600-2500
m (Howell and Webb 1995), which includes elevations similar to those inhabited
by relicta.
Paynter
(1970) recognized two species: Amaurospiza
concolor (relicta, concolor, aequatorialis) and A. moesta.
Lentino
and Restall (2003) suggested that based on wing formula, plumage and
morphological differences, and geographic distribution, aequatorialis could be a distinct species from concolor (Figure 5).
Notes
from HBW-BL: “plumage and mensural
differences are all minor, with the possible exception of the larger bill of carrizalensis;
moreover, new records from Brazil as far N as Maranhão suggest that populations
of Amaurospiza may generally be more widespread and less disjunct
than range maps indicate, as seems often the case with bamboo specialists.“

Figure 5. Comparative wing formulae of Amaurospiza as described by Lentino and
Restall (2003).
Howell
and Dyer (2022) commented on the similarity in morphology and voice of the taxa
within the genus Amaurospiza.
However, they noted that relicta is a
distinctive taxon, endemic to Mexico, and that it has been considered a
separate species. They also considered concolor
and aequatorialis to be conspecific.
New information:
Genetics
Bryson
et al. (2014) studied the diversification of the “blue cardinals” across the
New World. They generated multilocus sequence data from one mitochondrial gene
(ND2) and three nuclear introns (ACO1, MYC, FGB-I5) and estimated
time-calibrated species trees. The authors included four Amaurospiza taxa, all except for relicta. The mtDNA phylogeny recovered two main clades: a first
clade consisting of concolor (concolor) from southern Mexico and
Central America, and a second clade formed by the South American subspecies of concolor (aequatorialis), moesta,
and carrizalensis (Figure 6).
Therefore, Amaurospiza concolor was
not recovered as a monophyletic taxon, although only one sample from concolor aequatorialis was included, and
as noted before, concolor relicta was
not included. Support for the node uniting moesta
and carrizalensis was a middling PP
≥0.70, whereas support for all others was ≥0.95.
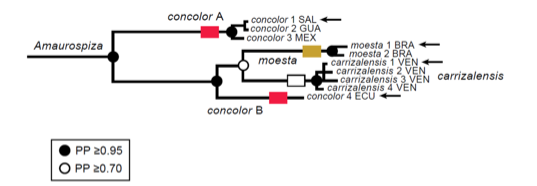
Figure 6. Relevant part of Figure 2 of Bryson
et al. (2014), mitochondrial ND2 Bayesian phylogeny.
The
multilocus phylogeny from Bryson et al. (2014) included a smaller sample size,
one individual per Amaurospiza taxa: concolor A (concolor), concolor B (aequatorialis), moesta, and carrizalensis.
However, the authors noted that it was not possible to obtain any nuclear data
for concolor B (aequatorialis), and this individual was represented only by mtDNA.
The divergence between Central American concolor
and South American aequatorialis, moesta, and carrizalensis was supported by a PP ≥0.95 (Figure 7). The nodes
within the South American clade had lower support (PP ≥0.70). Branch lengths in
Amaurospiza were comparable to
intraspecific divergence in Cyanocompsa
parellina and C. cyanoides,
although branch length between P. ciris
and P. versicolor was shorter, and
the branch length between Cyanoloxia and
“Cyanocompsa” brissonii was
comparable to Amaurospiza.
Genetic
evidence, based solely on mitochondrial DNA, suggests that concolor is more distantly related to aequatorialis than the latter is to carrizalensis and moesta;
therefore, the authors suggested that the geographically and genetically
distinctive aequatorialis be elevated
to species status (Bryson et al. 2014).
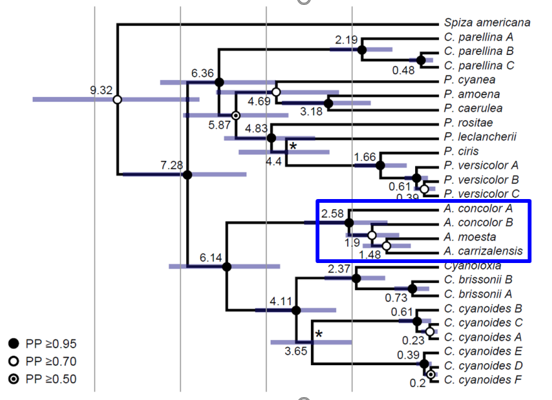
Figure 7. Multilocus *BEAST phylogeny from
Bryson et al. (2014).
The IOC list split aequatorialis
based on Bryson et al. (2014)
The SACC
assessed the split of aequatorialis
in 2016, analyzing the new phylogenetic information from Bryson et al. The
split was rejected mainly because vocal data were not published (Proposal 728, https://www.museum.lsu.edu/~Remsen/SACCprop728.htm).
Areta
et al. (2023) developed the first phylogenetic analysis that included multiple
samples from each of the five taxa within the genus Amaurospiza. The mitochondrial gene ND2 was sequenced for all 19
ingroup samples, and three nuclear introns (ACO1, FGB5, MB) for a subset of
samples (one sample per taxon, with the exception of relicta). ND2 and multilocus phylogenetic analyses confirmed the
monophyly of the genus Amaurospiza,
recovered A. moesta and A. carrizalensis as sister species, and
supported the relationship of aequatorialis
as sister to the moesta-carrizalensis clade, thus confirming the
paraphyly of A. concolor (Figure 8).
ND2 haplotypes of relicta were
recovered as monophyletic, either within a polytomy of concolor haplotypes in the ND2 gene tree or as sister to concolor in the BEAST tree. The
relationship of concolor + relicta (ND2) or concolor (multilocus) was recovered as sister to all the other
taxa.
Additionally,
Areta et al. (2023) estimated mean ND2 pairwise distances, showing that the
distances between concolor and aequatorialis were greater (8.3%) than
those between moesta and carrizalensis (5.7%). The two relicta samples diverged on average by
1.0% from nominate concolor.
Importantly, the authors uncovered low levels of intraspecific genetic
differentiation between geographically distant populations, which contrasts
with the deep divergences between allopatric species. Divergence times
estimated from ND2 suggest that the Central and South American groups diverged
6.1 Ma, that populations of relicta
diverged from concolor about 1
million years ago, and that the differentiation of South American lineages
started about 3.4 Ma (Figure 9).
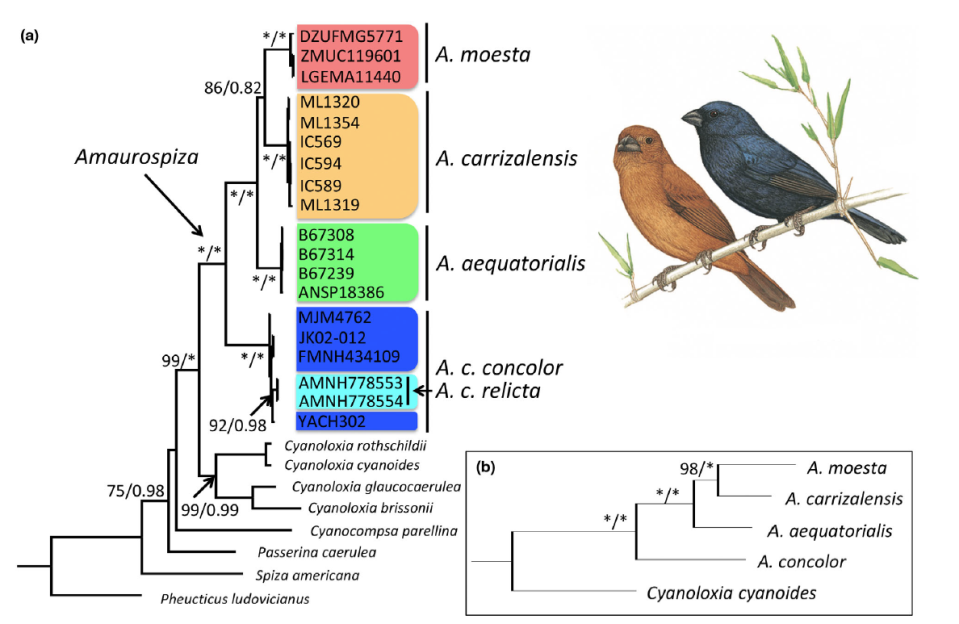
Figure 8. Phylogenetic hypothesis of
relationships within the genus Amaurospiza
from Areta et al. (2023). (a) mtDNA and (b) multilocus datasets. Numbers on
nodes represent maximum likelihood bootstrap (* 100%) / Bayesian posterior
probabilities (* 1.0).
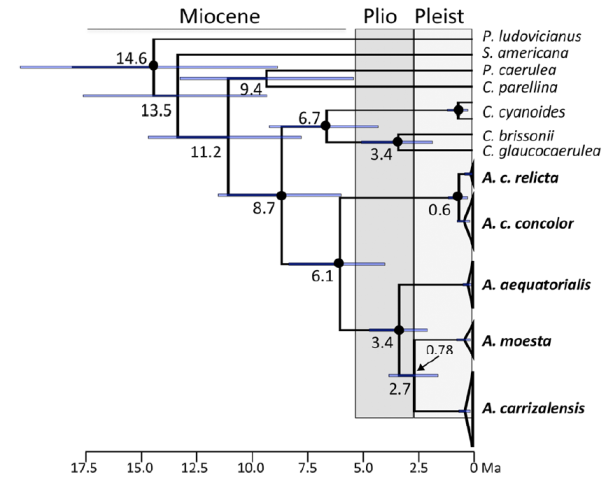
Figure 9. Bayesian phylogenetic reconstruction
of Amaurospiza based on ND2 data from
Areta et al. (2023).
Vocalizations
Boesman
(2016), using songs available in Xeno Canto (XC), analyzed and compared the
voices of concolor (including concolor and aequatorialis), moesta,
and carrizalensis. The taxon relicta was not included; there are no
songs available in XC or the Macaulay Library (only calls in XC). Boesman
concluded that the “song of all three species is very similar, given the range
of variation within each species”. He added:
“All basic sound parameters have a
largely overlapping range (min. frequency, max. frequency, number of notes,
note length, phrase length,...). Note shapes are also quite similar, with many
about identical between species.
“Other features that may allow differentiation such as e.g.
at start or end of a song phrase could not be found.
“It is probably impossible
to assign any recording with a reasonable level of confidence to any species. A
multivariate statistical analysis may allow to separate song of the different
taxa (once more recordings become available), but in any case differences will
be small, and will not lead to scores higher than e.g. 1 + 1 applying Tobias
criteria.”
Sample of the sonograms included by
Boesman (2016):
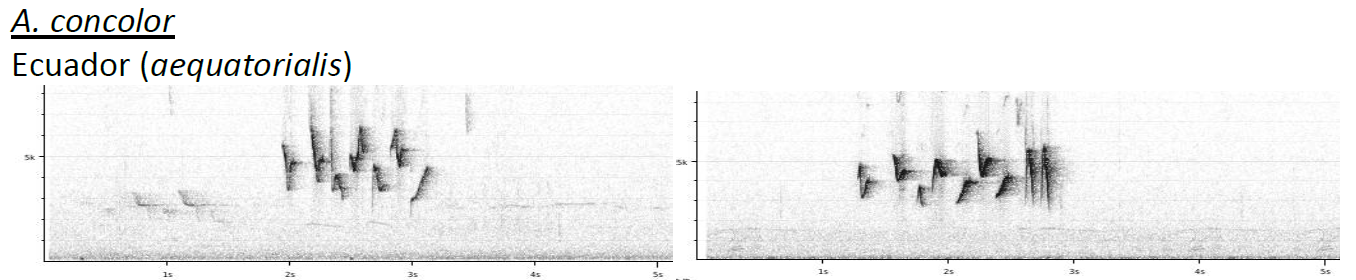
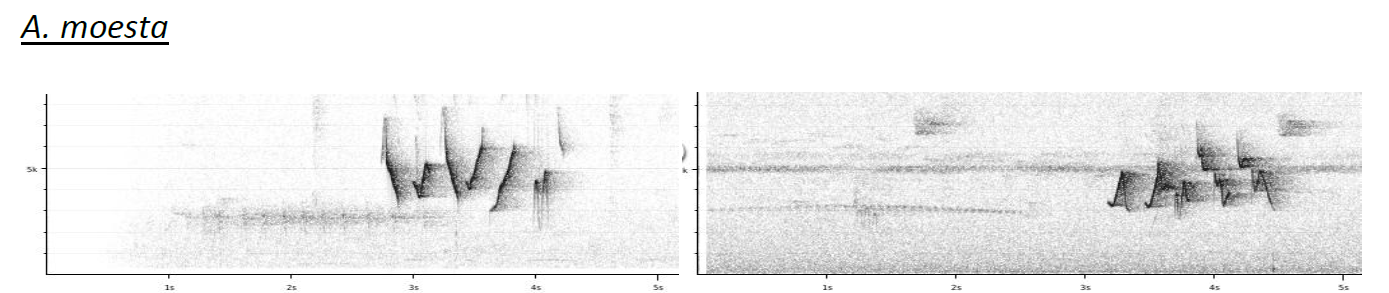
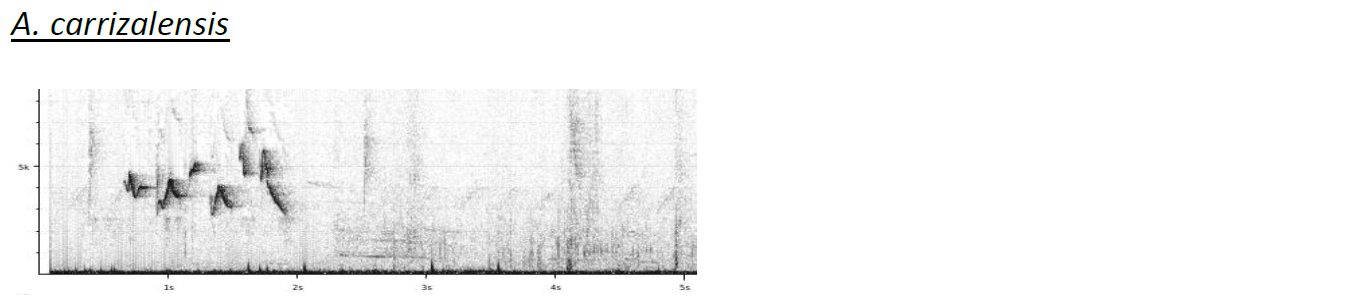
Notes from HBW-BL: “while relicta is here separated
as a full species, the other taxa appear to be very weakly differentiated:
available acoustic evidence reveals identical songs (Boesman 2016).”
Areta
et al. (2023) performed a quantitative vocal analysis that included the five
taxa within the genus Amaurospiza.
They showed that vocalizations are quite conserved in the group, but that they
also provide taxonomically useful information. The authors found consistent
differences between the Central and the South American clades: the number of
inflections/second exhibited a stepped pattern, with concolor and relicta on
the lower end and carrizalensis, aequatorialis, and moesta on the upper end; the South American taxa averaged more
inflections per note than concolor
and relicta (Table 6). A linear
discriminant analysis using nine acoustic variables correctly assigned all 62
songs to the correct taxon (but note that there were single recordings for relicta [most similar to nominate concolor] and carrizalensis [most similar to moesta].
The first linear discriminant consisted mainly of maximum frequency, peak
frequency average of all notes per song, and song duration on the first three
notes; this first linear discriminant separated the South American taxa from
the Central American taxa (Figure 10).
Table 6. From Areta et al. (2023), quantitative
characterization of the inflections in the songs of Amaurospiza seedeaters.
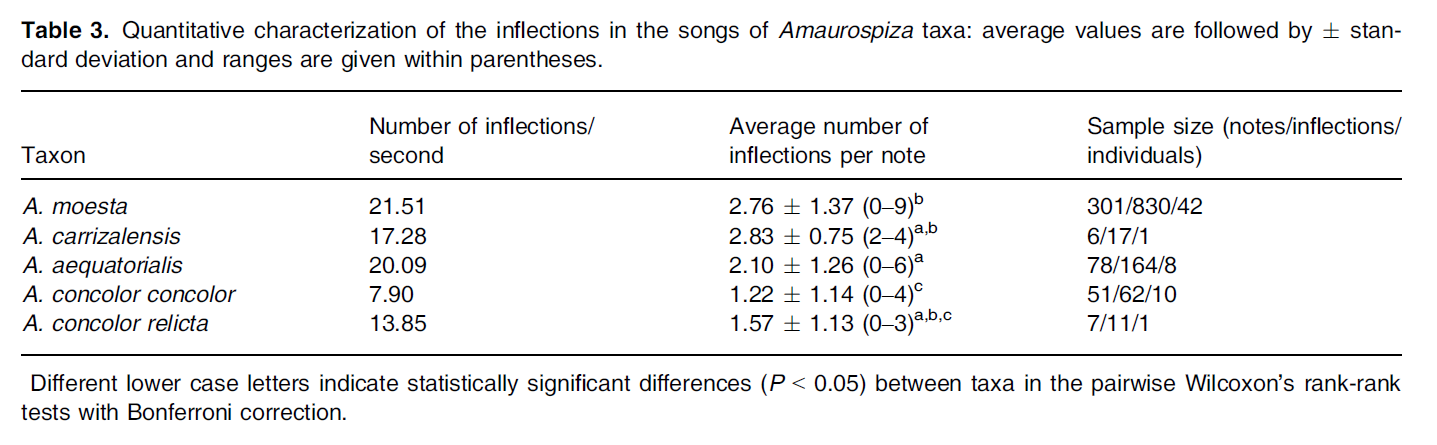
Figure 10. Linear discriminant analysis of
songs of Amaurospiza seedeaters from
Areta et al. (2023). Note the distinctive cluster of aequatorialis, and the placement of the single recordings of relicta (close to nominate concolor) and carrizalensis (close to moesta).
Recommendation:
Species
limits in Amaurospiza seedeaters are
a complex issue mainly due to their morphological similarity and allopatric
distributions. Each of the five Amaurospiza
taxa is considered a subspecies in at least one of the four global avian
checklists, and each has also been considered a separate species at some point
in history. Total evidence should be considered to reconcile the taxonomy of
these seedeaters. They all are allopatric with no evidence of intergradation,
show morphological differences, have similar but distinctive songs, and are
phylogenetically closely related. The recent integrative study by Areta et al.
(2023), which analyzed phylogenetic data, vocalizations, morphology, and
plumage, suggested that four species should be recognized within the genus Amaurospiza: A. concolor (relicta + concolor), A. aequatorialis, A.
carrizalensis, and A. moesta.
We
present four separate subproposals to revise the taxonomy of Amaurospiza seedeaters:
(a)
Split Amaurospiza
relicta from Blue Seedeater A.
concolor.
(b)
Lump two
subspecies of A. concolor (concolor
+ aequatorialis) and A. carrizalensis with A. moesta.
(c)
Split A.
aequatorialis from A. concolor.
(d)
Lump the five taxa (relicta, concolor, aequatorialis, carrizalensis, moesta) as
subspecies of Amaurospiza moesta.
Approval
of subproposals (a) and (b) would reconcile NACC (and SACC) with HBW-BL.
Approval of subproposal (c) would reconcile NACC (and SACC) with IOC and
follows the recommendation by Areta et al. (2023). Approval of (a) and (c)
would result in five species; conversely, approval of subproposal (d) would
lump the five taxa in a single species, A.
moesta.
We
recommend the following votes:
(a)
NO, different lines of evidence (genetics,
plumage, morphology, and vocalization) suggest that relicta should not be given species status but considered a
subspecies of A. concolor. However,
Areta et al. 2023 recommend more rigorous studies on the taxonomic status of relicta.
(b)
NO, neither Bryson et al. (2014) nor Areta et
al. (2023) provided phylogenetic support for this lump. HBW-BL considers concolor, aequatorialis, carrizalensis,
and moesta as subspecies within A. moesta, leaving A. relicta as a separate species. However, the two large clades (relicta, concolor / aequatorialis,
carrizalensis, moesta) supported by phylogenetic data do not correspond with that
classification.
(c)
YES, all evidence support aequatorialis as a separate species from A. concolor, aequatorialis
is more closely related to carrizalensis
and moesta than to concolor, and also differs from the
latter in having white underwing coverts (at least in adult males) and in song.
Considering aequatorialis as a
separate species requires a change in the geographic distribution of A. concolor to include only the area
from Mexico to Panama (eliminating Colombia and Ecuador).
(d)
NO, phenotypic and genotypic data do not
support the lump of the five taxa within a single species.
English names:
Similarly
to taxonomic treatment for Amaurospiza
seedeaters, there is no consensus in English names among global avian
checklists (Table 7).
Table
7. English names currently used for Amaurospiza
seedeaters in four global avian checklists.
|
|
Howard & Moore |
eBird/Clements |
HBW-BL |
IOC |
|
relicta |
Blue
Seedeater |
Blue
Seedeater (Slate-blue) |
Slate-blue
Seedeater |
Cabanis's Seedeater |
|
concolor |
Blue
Seedeater |
Blue
Seedeater (Blue) |
Blue
Seedeater |
Cabanis's Seedeater |
|
aequatorialis |
Blue
Seedeater |
Blue
Seedeater (Equatorial) |
Blue
Seedeater |
Ecuadorian Seedeater |
|
carrizalensis |
Carrizal
Seedeater |
Carrizal
Seedeater |
Blue
Seedeater |
Carrizal
Seedeater |
|
moesta |
Blackish-blue
Seedeater |
Blackish-blue
Seedeater |
Blue
Seedeater |
Blackish-blue
Seedeater |
The
NACC proposal reads as follows: “Therefore, according to passing subproposals,
please consider the following:
“•
If (a) passes and relicta is
separated, Slate-blue Seedeater could be used.
“•
If (b) passes and concolor (concolor + aequatorialis), carrizalensis,
and moesta are merged, the name Blue
Seedeater could continue to be used or a new name could be proposed.
“•
If (c) passes and aequatorialis is
separated, Areta et al. (2023) suggested the English name Ecuadorian Seedeater
because most of its range occurs in Ecuador, whereas the previously proposed
name, Equatorial Seedeater, could suggest a lowland distribution rather than
the montane range that the species occupies. If you vote YES on (c), please
vote either for Ecuadorian Seedeater or Equatorial Seedeater.
“•
If (d) passes and the five taxa become subspecies within A. moesta, a new English name should be proposed.
“•
If (a) and/or (c) pass, the name Blue Seedeater could continue to be used for concolor or a new name could be
proposed.
[Note
from Remsen to SACC voters: no need to vote now, but preliminary opinions
welcomed, A separate proposal that includes all the members of our English
names subgroup is a better way to go once we have voted on species limits.]
Literature cited:
Angulo Pratolongo, F.,
J. N. M. Flanagan, W. P. Vellinga, and N. Durand. (2012). Notes on the birds of
Laquipampa Wildlife Refuge, Lambayeque, Peru. Bulletin of the British
Ornithologists’ Club 132(3):162–174.
American
Ornithologists' Union. (1983). Check-list of North American birds. 6th edition.
American
Ornithologists' Union. (1998). Check-list of North American birds. 7th edition.
Areta, J. I., Benítez
Saldívar, M. J., Lentino, M., Miranda, J., Ferreira, M., Klicka, J., &
Pérez‐Emán, J. (2023?)
Phylogenetic relationships and systematics of the bamboo‐specialist
Amaurospiza blue‐seedeaters.
Ibis (in press).
Boesman, P. (2016).
Notes on the vocalizations of Blackish-blue Seedeater (Amaurospiza moesta). HBW
Alive Ornithological Note 388. In: Handbook of the Birds of the World Alive.
Lynx Edicions, Barcelona. https://doi.org/10.2173/bow-on.100388
Bryson Jr, R. W.,
Chaves, J., Smith, B. T., Miller, M. J., Winker, K., Pérez‐Emán,
J. L., & Klicka, J. (2014). Diversification across the New World within the
‘blue’ cardinalids (Aves: Cardinalidae). Journal of Biogeography, 41(3),
587-599.
Cabanis, J. 1861. Uebersicht der im Berliner Museum befindlichen Vögel von Costa Rica, Vom
Herausgeber. Journal für Ornithologie 9: 1-11.
Davis, L. I. (1972). A
Field Guide to the Birds of Mexico and Central America. University of Texas
Press, Austin.
Eisenmann, E. (1955).
The Species of Middle American Birds. Transactions of the Linnaean Society,
Volume VII, New York.
Griscom, L. (1934). The
ornithology of Guerrero, Mexico. Bull. Mus. Comp. Zool. 75: 367–422.
Hellmayr, C.E. 1938.
Catalogue of birds of the Americas. Part XI. Field Museum of Natural History
Zoological Series, volume 13, part 11.
Howell, S. N. G., and
S. Webb (1995). A Guide to the Birds of Mexico and Northern Central America.
Oxford University Press, New York, NY, USA.
Lentino, M. and
Restall, R. (2003). A new species of Amaurospiza blue seedeater from
Venezuela. Auk 12115(3): 6115115–61156.
Lopes, L. E., De Pinho,
J. B., & Benfica, C. E. R. (2011). Seasonal distribution and range of the
Blackish-blue Seedeater (Amaurospiza moesta): a bamboo-associated bird.
The Wilson Journal of Ornithology, 123(4), 797-802.
Miller, A.H., H.
Friedmann, L. Griscom, and R.T. Moore. 1957. Distributional check-list of the
birds of Mexico. Part 2. Pacific Coast Avifauna number 33.
Monroe, B. L., Jr.
(1968). A Distributional Survey of the Birds of Honduras. Ornithological
Monographs 7. American Ornithologists’ Union, Washington, DC, USA. 458 pp.
Orr, R.T., and M.S.
Ray. 1945. Critical comments on seed-eaters of the genus Amaurospiza.
Condor 47: 225-228.
Pablo-Castillo, J.
(2018). Nuevos aportes a la ecología e historia natural del semillero
azulado (Amaurospiza concolor, Cardinalidae) en Costa Rica. Comité
editorial.
Paynter, R. A., Jr.,
Ed. (1970). Checklist of Birds of the World, vol. 13. Museum of Comparative
Zoology, Harvard University Cambridge, Massachusetts.
Ramos-Ordoñez, M. F.,
C. I. Rodríguez-Flores, C. A. Soberanes-González, M. d. C. Arizmendi, A.
Jaramillo, and T. S. Schulenberg (2020). Blue Seedeater (Amaurospiza
concolor), version 1.0. In Birds of the World (T. S. Schulenberg, Editor).
Cornell Lab of Ornithology, Ithaca, NY, USA. https://doi.org/10.2173/bow.blusee1.01
Ridgway, R. 1901. The
birds of North and Middle America. Part I. United States National Museum
Bulletin 50.
Rising, J. D., A.
Jaramillo, G. M. Kirwan, and E. de Juana (2020). Blackish-blue Seedeater (Amaurospiza
moesta), version 1.0. In Birds of the World (S. M. Billerman, B. K. Keeney,
P. G. Rodewald, and T. S. Schulenberg, Editors). Cornell Lab of Ornithology,
Ithaca, NY, USA. https://doi.org/10.2173/bow.blbsee3.01
Sánchez, C., J. R.
Saucier, P. M. Benham, D. F. Lane, R. E. Gibbons, T. Valqui, S. A. Figueroa, C.
J. Schmitt, C. Sánchez, B. K. Schmidt, C. M. Milensky, A. García Bravo, and D.
García Olaechea. (2012). New and noteworthy records from northwestern Peru, Department
of Tumbes. Boletín de la Unión de Ornitólogos del Perú 7(2): 18–36.
Sharpe, R.B. 1888.
Catalogue of the birds in the British Museum. Volume XII. British Museum
(Natural History), London.
Rosa Alicia Jiménez, Terry Chesser and Juan I. Areta,
February 2023
Comments from Remsen: “Yes to the proposal’s recommendations, i.e. YES to “(c)” and NO to
a-b-d. But only because this seems to be
the best overall taxonomy given the collection of weak data we have and the
lack of stability in treatment of these taxa in various classifications. The proposal does a terrific job of
assembling every bit of published information on the group, but even combining
all this information, I see big problems.
The voices may be separable in DFA space, are those differences really
significant? DFA on the geographic
dialects of a related species, Passerina cyanea, would likely be able to
discriminate them as well. I lean more
towards Boesman’s qualitative impression that the songs are very similar,
remarkably so in my opinion given the great distances involved from one end of
this group to the other and the gaps in distribution. Playback trials would seem to be a necessary
step The genetic data are of interest, of course, but are weak in terms of
sampling of genes (mostly ND2), taxa, N, and geographic sampling with
populations. That’s not the authors’
fault – these are mostly hard-to-find, rare-to-uncommon birds – the studies did
the best they could with the material available. The minor differences in bill size, body
size, wing formulas, and coloration are all matched by intraspecific variation
in these characters in broadly distributed polytypic species of other
passerines and in themselves seem only to confirm that the taxa are
phenotypically diagnosable and thus worthy of at least subspecies rank under
the BSC. Nonetheless, some of these
differences are roughly comparable to phenotypic differences in related taxa we
rank as species, specifically Cyanoloxia cyanoides/C. rothschildii,
for which we have better data.”
Comments from Areta: “YES to C,
given the closer relationship of
aequatorialis to South
American taxa evidenced in both mitochondrial and nuclear markers, the plumage
differences, and the vocal distinctions.
NO to A, B and D. The songs of
Amaurospiza are rather
conserved, but not identical among taxa (note that there is just one recording
for carrizalensis
and one for relicta, that in
our analyses were more similar to their closest relatives). However, note that
relicta is
apparently very recently diverged from nominate
concolor (mtDNA only),
suggesting that for the time being and until there is more data, the current
evidence supports retaining it as a ssp. of
concolor. I don´t see any evidence supporting the merger of any other
taxon with moesta.”
Comments from Robbins: “Kudos to the authors of this
proposal for pulling all the information together in a digestible format. With
some hesitation, I vote YES to subproposal C, for recognizing aequatorialis
as a species, and NO to the other subproposals.
“As Van pointed out, there are issues with saying anything
meaningful about vocalizations beyond that they are very similar. Similarly,
plumage and morphometrics don’t offer much insight into this taxonomically
difficult complex. As we increasingly see, phylogenetics based solely on a
single mitochondrial gene can be misleading, but given the rather large mean
pairwise distances between aequatorialis and concolor, in
comparison to related taxa, leads me to put more weight on that factor than any
other. Hence, my vote to recognize aequatorialis as a species.”
Comments from Stiles: “The genetic data favor at least the
split of Mesoamerican concolor from South American aequatorialis,
carrizalensis and moesta and the vocal data clearly separate aequatorialis
from moesta; the separation of carrizalensis from moesta
is less clear, but given that SACC recognized it, I’ll tentatively continue to
do so as well. So, (b) NO, (c) YES, (d) NO.”
Comments from Lane: A) NO, B)
NO, C) YES, D) NO. This result seems best to reflect the molecular and voice
data presented in the proposal. These taxa are all quite rare and difficult to
document in my limited experience, so it will be a challenge to try to augment
the dataset here to see if it will change the results. I do think “Ecuadorian
Seedeater” would be an appropriate name for
A. aequatorialis. Luckily, dealing with
relicta is not our
burden to bear.”
Comments from Bonaccorso:
“969a. NO.
The data from Areta et al (2023) do not show much of a genetic difference between
A. c. concolor and A. c. relicta. Given that “The song of relicta
is described as similar to concolor but slightly higher and faster
(Howell and Webb 1995),” I don’t see much of a strong argument to split them.
Minor bill shape, size, and color differences may be just as those expected
among subspecies.
“969b. NO.
I don´t see how lumping A. moesta and A. carrizalensis makes
sense; they show almost the same amount of genetic differentiation seen between
A. moesta + A. carrizalensis and A. aequatorialis. Because
morphological differences are overlapping and song data for carrizalensis
in the LD1 is so scant, we need to go with the phylogenetic data.
“969c. YES.
Clearly, A. aequatorialis and A. concolor are not monophyletic.
So, splitting A. aequatorialis from A. concolor is the sensible
option.
“969d. NO.
For the reasons stated in the proposal (phenotypic and genotypic data do not
support the lump of the five taxa).”
Comments
from Claramunt: “YES to (c) treat A.
aequatorialis as a
separate species from A.
concolor. This is the
important piece of evidence provided by the molecular phylogeny, as pointed out
in the previous proposal 728: A. aequatorialis is not sister to concolor but to the other South American clade, and is
genetically and phenotypically (maybe in subtle ways) differentiated from all.”
“NO to
a,b,d. No evidence supports these changes.”
Comments
from Zimmer:
“Revise the taxonomy of Amaurospiza seedeaters. This is, as several others have already
pointed out, a tough one, given that sample sizes for genetic material and
vocal material are very limited, and, in general, this entire group is pretty
poorly known. But here goes…
A)
“NO.
Range disjunctions, non-overlapping morphometric characters, and
possible subtle distinctions in vocalizations all point to continued
recognition of relicta as a distinct subspecies of concolor, but
lacking genetic data corroborating greater divergence and a better sampled,
quantitative vocal analysis, I don’t think there’s enough here to support a
split.
B)
“NO
to lumping concolor + aequatorialis and carrizalensis into
moesta. This runs counter to the
genetic data, my impression of vocal differences, and ecological and
biogeographic considerations.
C)
“YES
to splitting aequatorialis from concolor, based on genetic data
(demonstrating paraphyly of concolor if aequatorialis is
included, and, the sister relationship of aequatorialis to the moesta-carrizalensis
clade), differences in underwing covert color in adult males (seemingly an
important signaling mechanism, based on how conspicuous the white underwing
“flash” is in the field for those “seedeaters” that have it), apparent
differences (no matter how subtle) in vocalizations, and biogeographic and
ecological considerations.
D)
“NO. Morphological/vocal data, genetic data, and
ecological distinctions fail to provide support for lumping the five taxa, in
my opinion.”