Proposal (979)
to South
American Classification Committee
Revise the taxonomy of
the Charadriidae (in 8 parts, A-H)
Background:
The
Charadriidae comprises eighteen species in two subfamilies in the SACC area. Of
these, ten are in the long-recognized genus of cosmopolitan Charadrius plovers.
The current linear sequence (Remsen et al.,
2023) for the species in this family is:
Pluvialis dominica American Golden-Plover
Pluvialis fulva Pacific Golden-Plover
Pluvialis squatarola Black-bellied Plover
Oreopholus ruficollis Tawny-throated
Dotterel
Vanellus cayanus Pied Lapwing
Vanellus chilensis Southern Lapwing
Vanellus resplendens Andean Lapwing
Charadrius modestus Rufous-chested
Dotterel
Charadrius vociferus Killdeer
Charadrius semipalmatus Semipalmated Plover
Charadrius melodus Piping Plover
Charadrius mongolus Lesser Sand-Plover
Charadrius wilsonia Wilson's Plover
Charadrius collaris Collared Plover
Charadrius alticola Puna Plover
Charadrius falklandicus Two-banded Plover
Charadrius nivosus Snowy Plover
Phegornis mitchellii Diademed
Sandpiper-Plover
New
Information:
A
series of papers has come out addressing the phylogenetic relationships in the
Charadriidae, with increasing amounts of data. However, all papers suffer from
having few genetic markers, which hampers our ability to confidently make
changes to genera, and two papers additionally suffer from limited taxon
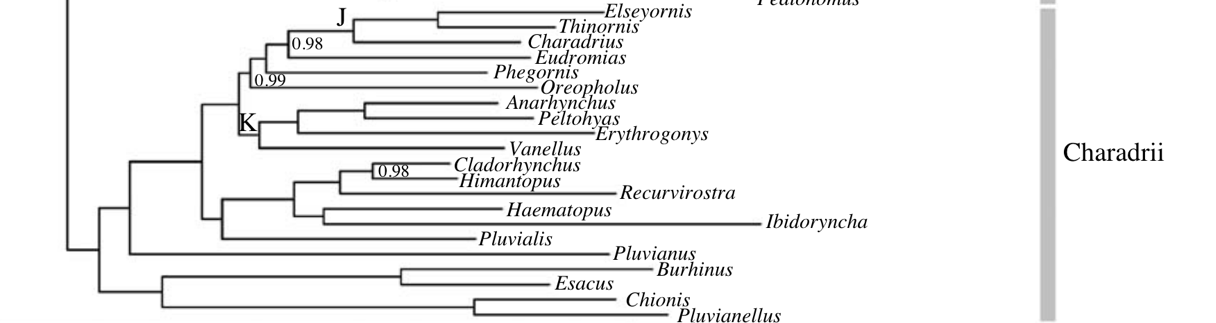
sampling. Baker et al. (2007) sampled
three mitochondrial loci and one nuclear locus, targeting one sample per genus.
Below is the relevant part of their tree. Note that Anarhynchus is in
the same clade as Vanellus, the two of which are unrelated to the one Charadrius
sample. Nodes without support values have a posterior value of 1. This study,
and one a few years later (Baker et al. 2012) were the basis of SACC
proposal 551,
which rearranged the linear sequence of Charadriiform genera, but not genus
limits.
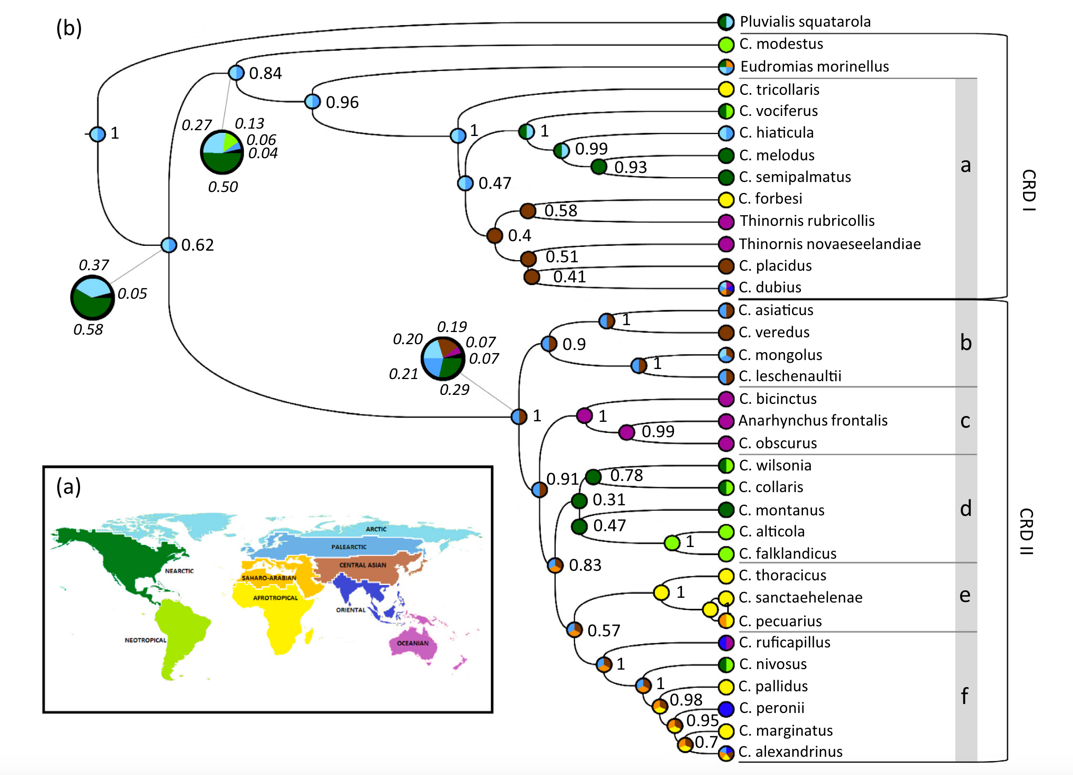
Dos Remedios et al. (2015) sampled two mitochondrial and four nuclear
loci from 29 Charadrius species. Their Bayesian consensus tree is shown
below. Node values are posterior probabilities. Note a lack of any Vanellus
samples, and the low node support in many parts of the tree. This phylogeny was
the basis of SACC proposal 735-F, which rearranged the
linear sequence of species in Charadrius but did not change genus
limits. The map and colored circles are a biogeographic analysis.
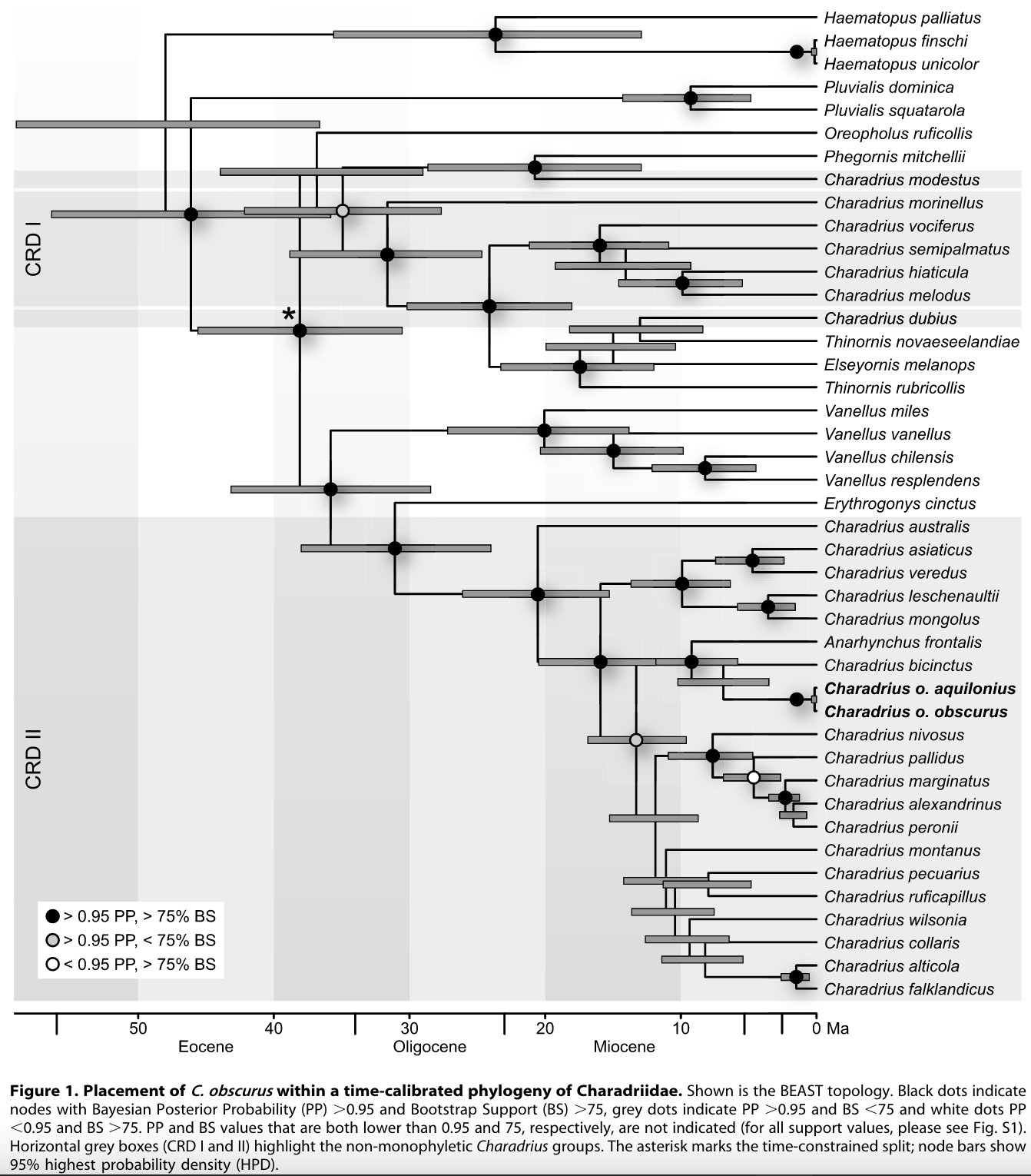
Another
study (Barth et al. 2013) sampled eight mitochondrial and two nuclear loci from
a decent selection of Charadriidae but focusing on placing the New Zealand
Dotterel (Charadrius obscurus) in the phylogeny. Their results largely
corroborate those of the other papers outlined here. Their Figure 1 phylogeny
is shown below. Note that nodes without circles have low support and should be
considered polytomies.
Lastly,
we come to the paper that is the basis for most of the changes in this
proposal. Černý & Natale (2022) used a supertree approach that utilized
much of the same data as in the previous papers (Baker et al. 2007, Barth et
al. 2013, Dos Remedios et al. 2015), with the addition of more loci (27 total)
and a matrix of 69 skeletal characters, to address the relationships of most
species in the order Charadriiformes. The main issue I have with this paper is
that the main figure (screenshot below) combines both the genetic and skeletal
data with no indication of whether the relationships are supported by one or
the other data types. I’m very skeptical of the older relationships in the main
figure, given well known issues of convergence in skeletal datasets. Nodes with
bootstrap support ≥70% are indicated by circles, and nodes with bootstrap
support <70% are indicated by squares. As you can see, there are many
squares on the nodes in this tree. This is no surprise, given the very high
percentage of missing data in the molecular data matrix (68.5%!) with some
samples represented by just a single locus (mean = 9.5 loci).
A portion of Figure 6 from Černý & Natale
(2022), showing phylogenetic relationships in the Charadriidae. Note that the
darker gray vertical lines are 10 Ma increments, and the faint gray lines are
2.5 Ma. Note that for converting this to a linear sequence, this tree should be
“read” bottom-to-top.
Thankfully,
the supplemental data for Černý & Natale (2022) does contain the
molecular-only phylogeny, but it shows that many of the relationships,
especially those deeper in the tree, are weakly supported by molecular data. To
better illustrate those relationships that do have good support in the tree, I
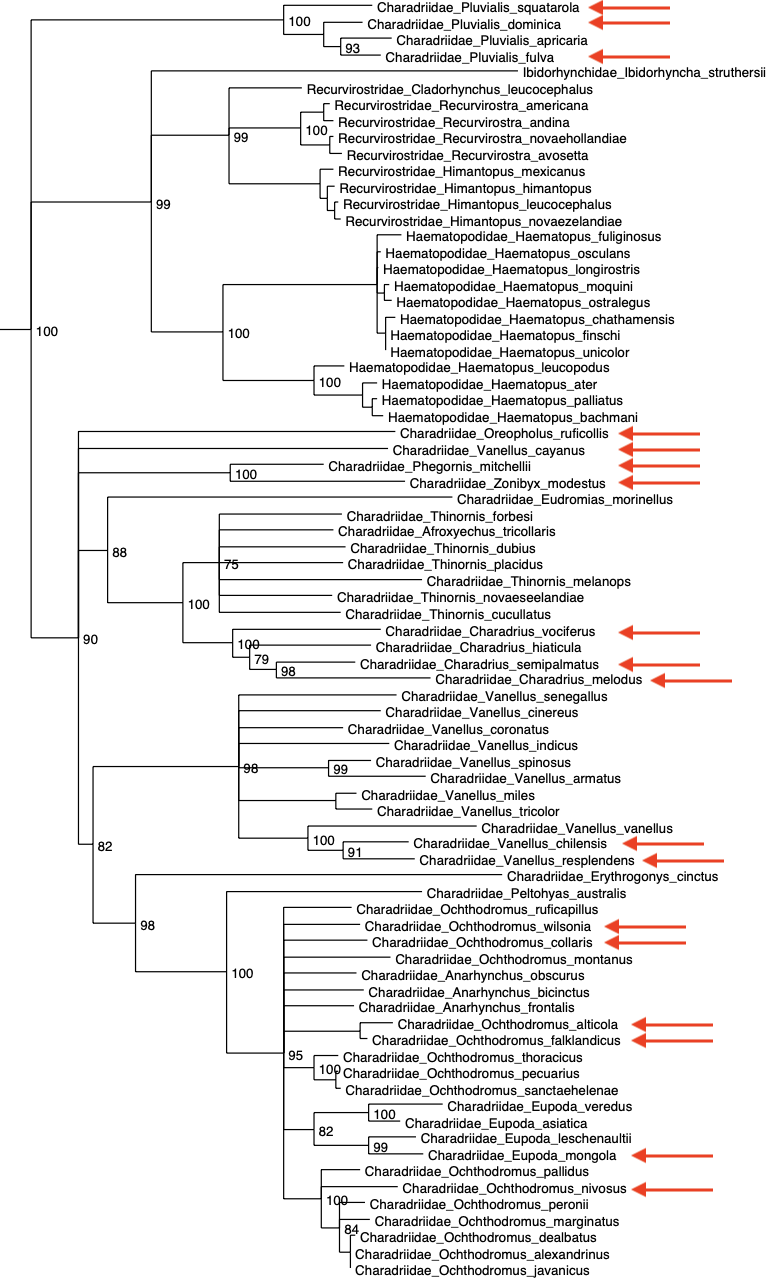
collapsed all nodes with posterior probabilities less than 0.75. Below is the
relevant portion of the tree, with the species occurring in the SACC area
indicated in red. Despite the weak support for many nodes, there is strong
support for four main clades; 1) Pluvialis, 2) Charadrius sensu
stricto, 3) Vanellus, and 4) the remainder of the Charadrius.
There are a handful of species on long branches, whose relationships within the
family are unresolved, including four species in the SACC area: Oreopholus
ruficollis, Vanellus cayanus, Phegornis mitchellii, and Charadrius
modestus. The relationship between Pluvialis and the rest of the
group, and especially whether it’s closer to the Recurvirostridae, is unresolved.
As
an aside, I’ve looked through the rest of this Charadriiformes tree and found
just one other case of genus-level paraphyly (based on current SACC taxonomy).
That topology places Sterna forsteri and S. trudeaui as
sister to Thalasseus plus the rest of Sterna. However, this
relationship is weakly supported, so I don’t think any changes should be
considered for now.
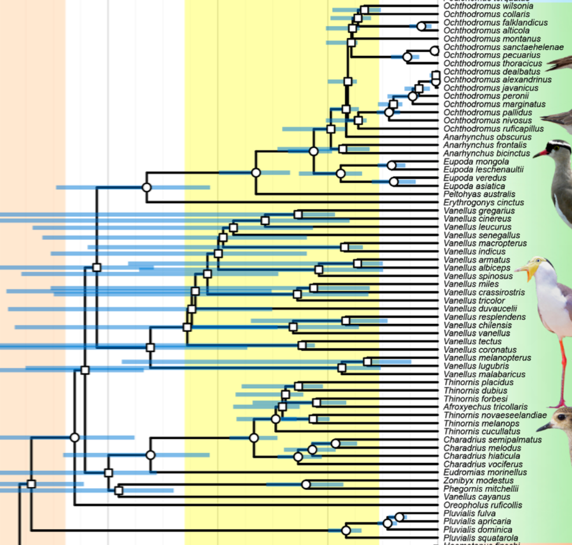
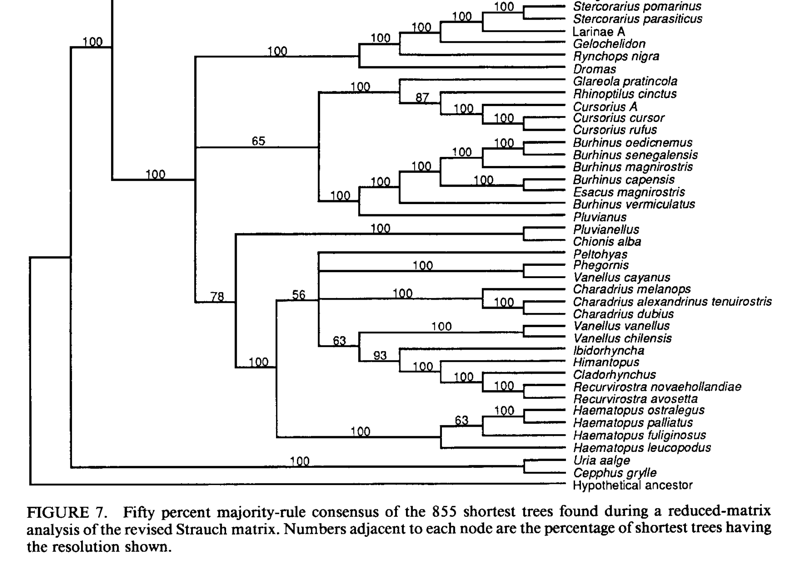
Out
of curiosity, I delved into the skeletal data used in this tree. Černý &
Natale (2022) used data from Strauch (1978),
which had been reanalyzed by Chu (1995).
Chu (1995) presented phylogenies based solely on the skeletal data, which I’ve
included below. As you can see, these deeper relationships strongly conflict
with the molecular data, and as expected, place unrelated taxa close to each
other in cases where morphology is similar (e.g. Vanellus closer to
Recurvirostridae in some trees). Another tree from Chu (1995; not shown) with
more tips and fewer markers, which includes more Charadrius tips, is
less well-resolved but places Vanellus in a big polytomy with Charadrius
sensu lato, so closer to where the molecular data indicate it belongs.
I
think there are a few options here. One would be to wait for better data, which
was my initial reaction on seeing the phylogeny of Černý & Natale (2022).
However, it seems that at least WGAC and Clements are moving forward with some
changes to genera, and I think it would be worthwhile for SACC to consider
following these global authorities and making some of these changes. Although
the molecular data are not resolving all the relationships in the group, they
do indicate that our current treatment of Charadrius is highly
paraphyletic, which would require a few changes. There are also a handful of
species (e.g. Vanellus cayanus) that are on long branches that suggest
that they could be split off as monotypic genera, even if their exact
relationships are currently unresolved or poorly supported.
To
make the taxonomy a bit easier to follow, I’ve listed here the author, year,
and type species for each of the genera that are relevant to this matter,
including those that could reasonably be resurrected for a clade. These are
sorted by year.
Charadrius Linnaeus 1758, type hiaticula
Vanellus
Brisson,
1760, type vanellus
Pluvialis
Brisson
1760, type apricaria
Anarhynchus Quoy & Gaimard
1830, type frontalis
Eudromias
Brehm
1831, type morinellus
Oreopholus, Jardine & Selby
1835, type ruficollis
Erythrogonys
Gould
1838, type cinctus
Thinornis Gray, GR 1844, type novaeseelandiae
Phegornis
Gray,
GR 1846, type mitchellii
Ochthodromus Reichenbach 1852, type
wilsonia
Zonibyx
Reichenbach
1852, type modestus
Eupoda Brandt, JF 1852, type asiatica
Hoploxypterus
Bonaparte
1856, type cayanus
Aegialophilus
Gould
1865, type alexandrinus
Peltohyas Sharpe 1896, type australis
Afroxyechus Mathews 1913, type tricollaris
Effect
on SACC area:
Following
the tentative recommendations below would lead to a new taxonomic arrangement
and linear sequence for the Charadriidae, as follows:
Pluvialis squatarola Black-bellied Plover
Pluvialis dominica American Golden-Plover
Pluvialis fulva Pacific Golden-Plover
Oreopholus ruficollis Tawny-throated
Dotterel
Hoploxypterus cayanus Pied Lapwing
Phegornis mitchellii Diademed
Sandpiper-Plover
Zonibyx modestus Rufous-chested
Dotterel
Charadrius vociferus Killdeer
Charadrius semipalmatus Semipalmated Plover
Charadrius melodus Piping Plover
Vanellus chilensis Southern Lapwing
Vanellus resplendens Andean Lapwing
Anarhynchus mongolus Lesser Sand-Plover
Anarhynchus wilsonia Wilson's Plover
Anarhynchus collaris Collared Plover
Anarhynchus alticola Puna Plover
Anarhynchus falklandicus Two-banded Plover
Anarhynchus nivosus Snowy Plover
Recommendation:
Please
vote on the following issues:
A.
Rearrange species in the genus Pluvialis
B.
Transfer Charadrius mongolus to Eupoda, and wilsonia, collaris, alticola, falklandicus, and nivosus to Ochthodromus
C.
Transfer Charadrius mongolus, wilsonia, collaris, alticola,
falklandicus, and
nivosus to Anarhynchus
D.
Adopt the new linear sequence
E.
Transfer Vanellus cayanus
to Hoploxypterus
F.
Transfer Charadrius modestus
to Zonibyx
G.
Transfer Charadrius modestus
to Phegornis
H.
Recognize the subfamily Pluvialinae for the species in Pluvialis, and the subfamily Charadriinae for
the remaining species in Charadriidae
Note
that both C & D and F & G are mutually exclusive.
I
recommend the following votes:
A.
YES: a minor bookkeeping change, supported by the internal relationships in Pluvialis.
B.
NO: despite my initial hesitation on making revisions in this group given the
haphazard quality of the phylogenetic data, there is strong evidence that this
clade of former Charadrius is unrelated to the core group, and is at
least as divergent as genera like Vanellus, so they could potentially go
in their own subfamily. Černý and Natale (2022) recommend recognizing Eupoda
but not Ochthodromus, but as shown in the phylogeny of molecular-only
data, there is very little support for the internal relationships in the group.
However, Barth et al. (2013) do also support this topology, with a somewhat old
(~17 Ma) split from the rest of this clade. I am including this one here for
sake of completeness, but also because there is a possibility that if the
topology in Černý and Natale (2022) holds up with better data, that it could be
a viable alternative. It doesn’t matter at all for the taxonomy, but the
etymology of Ochthodromus (“bank/shore-runner”) is much more appropriate
for this group of plovers than is Anarhynchus (“backward-bill”).
C.
YES. I highly recommend this option. The data are quite clear that this clade
should not be in Charadrius and given that we don’t have a good handle
on the internal topology, I think it’s safest to transfer all species to this
genus for the time being. This is also taxonomy that WGAC and Clements are
using, so it would put SACC in line with those global taxonomies. The one issue
that I foresee here is that there could potentially be an older genus than Anarhynchus available for one of the
species in this group. I did check the Richmond Index for the 28 synonyms of Charadrius
listed in Birds of the World and none superseded Anarhynchus. Frank
Rheindt and Pam Rasmussen have also done a thorough search for older available
names (WGAC comments) and are unable to find older available names.
D.
YES. Given the phylogenetic data that we have, this is the most accurate linear
sequence for the group. This mostly just moves Vanellus between the two
former Charadrius clades. I don’t think any changes should be made on
the linear sequence within each of the groups, given the low resolution in the
phylogenetic trees.
E.
YES. Vanellus cayanus is on a long branch that is much older than
many other groups considered genera (e.g. Vanellus
sensu stricto), sister to mitchellii + modestus. The Černý & Natale (2022)
phylogeny places the divergence time at about 28 Ma, which suggests genus-level
divergence. For what little it’s worth, the skeletal data in Chu (1995) also
place cayanus and mitchellii as sisters (modestus not included).
F.
YES. Charadrius modestus is on a long branch that is much older
than many other groups considered genera (e.g. Vanellus sensu stricto),
and is sister to Phegornis
mitchellii. The Černý &
Natale (2022) phylogeny places the divergence time at about 12 Ma. Given how
different these two species are, I think it’s better to keep these as separate
genera.
G.
NO. Just as an alternative to resurrecting Zonibyx, modestus
could be transferred to Phegornis. I don’t think that this is a great
option, given how distinctive these two species are, but the divergence time is
slightly less than the crown age of some of the other genera in the family.
H.
YES. I think it’s best to consider Pluvialis in a separate subfamily,
Pluvialinae. This clade is on a very long branch and could potentially even be
its own family.
Literature
Cited:
Baker, A. J., Pereira,
S. L., & Paton, T. A. (2007). Phylogenetic relationships and divergence
times of Charadriiformes genera: Multigene evidence for the Cretaceous origin
of at least 14 clades of shorebirds. Biology Letters, 3(2),
205–210. https://doi.org/10.1098/rsbl.2006.0606
Baker, A. J., Y.
Yatsenko, and E. S. Tavares. 2012. Eight independent nuclear genes support
monophyly of the plovers: the role of mutational variance in gene trees. Molecular Phylogenetics and Evolution 65:
631-641.
Barth, J. M. I.,
Matschiner, M. & Robertson, B. C. (2013). Phylogenetic position and
subspecies divergence of the endangered New Zealand Dotterel (Charadrius
obscurus). PLOS One, 8(10), e78068.
https://doi.org/10.1371/journal.pone.0078068
Černý, D., &
Natale, R. (2022). Comprehensive taxon sampling and vetted fossils help clarify
the time tree of shorebirds (Aves, Charadriiformes). Molecular Phylogenetics
and Evolution, 177, 107620.
https://doi.org/10.1016/j.ympev.2022.107620
Chu, P. C. (1995).
Phylogenetic reanalysis of Strauch's osteological data set for the
Charadriiformes. The Condor, 97(1), 174–196.
https://doi.org/10.2307/1368995
Dos Remedios, N., Lee,
P. L. M., Burke, T., Székely, T., & Küpper, C. (2015). North or south?
Phylogenetic and biogeographic origins of a globally distributed avian clade. Molecular
Phylogenetics and Evolution, 89, 151–159.
https://doi.org/10.1016/j.ympev.2015.04.010
Remsen, J. V., Jr., J.
I. Areta, E. Bonaccorso, S. Claramunt, G. Del-Rio, A. Jaramillo, D. F. Lane, M.
B. Robbins, F. G. Stiles, and K. J. Zimmer. Version 1 August 2023. A
classification of the bird species of South America. Museum of Natural Science,
Louisiana State University. http://www.museum.lsu.edu/~Remsen/SACCBaseline.htm
Strauch, J. G. (1978).
The phylogeny of the Charadriiformes (Aves): a new estimate using the method of
character compatibility analysis. The Transactions of the Zoological Society
of London, 34(3), 263–345.
Oscar Johnson, August
2023
Comments from Areta: “I vote in favor of all the options recommended by Oscar
(i.e., NO to B and G, and YES to the rest). I have always been in favor of Hoploxypterus, and it is also great to see Zonibyx coming back to
life. These genus-level changes were recently voted on in the working list of
the WGAC."
Comments from Robbins: “Oscar’s
recommendations are well reasoned and given that WGAC has gone that route,
makes sense to follow. I vote for all
suggestions Oscar recommended.”
Comments
from Zimmer: “A) YES; B) NO; C) YES; D) YES; E) YES – make
that a strong YES (Placing cayanus in Vanellus always bothered me
on several fronts, including structure, size, plumage, vocalizations, etc.); F)
YES; G) NO – make that a strong NO (Phegornis is truly a different
beast, and should be kept monotypic in my opinion.); H) YES – for all of the
reasons cited by Oscar in the Proposal.”
Comments from Lane:
“A. YES.
B. NO.
C. YES. But
I chuckle a little at this, as it takes a monotypic genus with the sole member,
Wrybill, defined by one of the bird world's more unusual bills, and expands it
to a rather speciose group!
D. YES.
E. YES.
F. YES.
G. NO.
H. YES.”
Comments
from Remsen: “YES on all of Oscar’s recommendations. Kudos to Oscar for extracting the reliable
signal out of all that messy data, and all the recommendations are cautious and
wise in my opinion. That the banded
plovers fall in two different genera is another example of plumage characters
not necessarily providing reliable phylogenetic signal. However, that morinellus is on a long
branch that argues for restoration of Eudromias is subjectively pleasing
to those of us who never understood its inclusion in Charadrius. In the NACC comments, Kevin Winker made a
good point on what we can/can’t make of a long branch when there are so many
missing data, but I think the new data shifted the balance in what is an
arbitrary decision in terms of strict typology.
“Mini-rant:
That the gene-based tree was not published as a figure in the Černý-Natale paper is
yet another of dozens of examples in the last decade that make me wonder
sometimes if MPE is actually a peer-reviewed journal. Evidently, the message is not widely
appreciated that phenotypic data have a difficult time eliminating convergence
or strong divergence, as “proven” by the Livezey-Zusi monograph that produced a
couple dozen conspicuous and indisputable examples of this (e.g., loons and
grebes maintained as sisters, and diving-petrels and phalaropes restored to
family rank). That was way back in 2007
with the world’s largest-ever matrix of phenotypic data (2K+ characters). I really like morphology and analyses of
morphological data. Morphology is the
phenotypic expression of all those genes on which natural selection can
act. Morphological data often map well
onto phylogeny but let’s see what the genes-only data look like for comparison. It’s the discrepancies that often provide
insights into the process of evolution.”
Comments
from Bonaccorso:
“A. NO. As
far as I can see, the Černý & Natale (2022) is the only phylogeny that
includes Pluvialis squatarola,
dominica, and fulva, and does
not have good support for the dominica
+ fulva
relationship.
“B. NO. I
might be wrong, but it seems to me that the proposal does not state clearly why
this would be the best arrangement.
“C. NO, for
the same reason stated in 979B.
“D. NO, as
it follows from my NO on proposals A-C.
“E. NO. I
understand that Hoploxypterus
(Latham, 1790) would be
available for cayanus
but what if cayanus
is also related to another species whose name could have priority
over Hoploxypterus? I think
it is too soon to make the change.
“F. NO. I
understand that Zonibyx
(Lichtenstein 1823) would be available for
modestus. It could go to either Zonibyx
or to Phegornis
“(979G), but I think these
decisions are premature given how fragmentary the phylogenetic evidence is.
“G. NO.
This decision is premature given the scarcity and patchiness of the
phylogenetic evidence.
“H.
YES. But Oreopholus
ruficollis is also in a very long
branch. Should we place it in its own subfamily too?”
Response
to Bonaccorso comments from Oscar Johnson: “I certainly agree that the phylogenetic data
that we have available are quite poor, and that makes it a real challenge to
confidently convert this to a genus-level taxonomy. I had initially advocated
(to NACC) that we basically ignore the Cerny and Natale paper for the time
being, given how many unresolved relationships there were in the tree, but
given that at least some global checklists were moving forward with some
changes, I felt it would be better to get ahead of those changes by
highlighting some of the very major issues in the underlying data, with the
hope that a more moderate decision by NACC or SACC might prevent a wholesale
adoption of the taxonomic recommendations advocated for in these papers. I do
think that some of the changes I advocate for in the proposal have reasonably
good support, but I fully endorse a more moderate/conservative decision if
that's how you or other committee members choose to proceed. Below are
some more details on each of the sub-proposals:
“A) The linear sequence rearrangement I presented for
Pluvialis moves
squatarola to the beginning of
the genus, which is supported in Cerny and Natale (2020) by a 100% bootstrap
support on the branch leading to squatarola, but does not change the
order of dominica
vs
fulva due to the poor resolution in this part of
the tree.
“B/C) The phylogenetic data in Cerny and Natale (2020) and prior
papers (especially Barth et al. 2013) all support this clade of former
Charadrius being more closely related to
Vanellus than to Charadrius sensu
stricto, thus rendering Charadrius paraphyletic and requiring a
change in genus limits if the phylogenetic data are to be trusted. I certainly
understand wanting to wait for better data given that these papers are all
based on relatively few loci, but the bulk of the evidence (to me) point
towards paraphyly in Charadrius. I gave these two
sub-proposals as competing options on how to circumscribe the genus limits in
this clade of former Charadrius, with the recommendation of adopting the
minimal number of new genera (Yes on C). A Yes on B would take this clade of
former Charadrius and adopt three new
genera: Anarhynchus,
Eupoda, and
Ochthodromus, which was what was advocated for by Cerny and Natale,
but as is clear from the molecular-only phylogeny in that paper, the
relationships within
this
clade are largely unresolved, so I recommended instead transferring all of
these species to Anarhynchus (Yes on C) and awaiting better phylogenetic
data before further splitting the genus.
“D) Other than re-arranging the position of the newly
circumscribed genera that I advocate for in the sub-proposals, I did not change
the linear sequence within any genera due to the largely unresolved
phylogenetic data. I could easily have made a mistake here, so it's worth
double-checking my work. Also, if some of the genus limits I recommend are not
adopted, then this linear sequence would need to be revised.
“E/F/G) These (potentially) monotypic genera are certainly the
toughest part of these sub-proposals, which is why I gave each one as a
separate voting option in case the committee wanted to accept/reject each one
individually. It's certainly possible that
Vanellus/Hoploxypterus cayanus is related to another clade, but
aside from Vanellus there's no other clade that it would be clearly
allied to in terms of morphology. In other words, even if it is sister to
another genus on a long branch, I'm not sure it would make sense to merge it
with that other genus based on its different morphology. Leaving it in
Vanellus would certainly be a valid option for the time being. For
modestus, I really don't know what the best solution is here, which is why
I gave two competing options. The phylogenetic data does seem to support it
being sister to mitchellii, however, rather than part of
Charadrius.
“H) It's certainly possible that
Oreopholus is on a long-enough branch to be its own subfamily, but
the phylogenetic data (such as it is) does indicate that it's in a polytomy at
the base of the rest of Charadriinae, so this seems like it would be premature.
Pluvialis, on the other hand, is in a polytomy at the base of the
node uniting not only the rest of Charadriidae, but also Recurvirostridae and
Haematopodidae, so moving it to its own subfamily seems to me to be the more
conservative option (versus its own family).
Additional
comments from Bonaccorso: “I vote in favor of all the options supported by Oscar
(NO to B and G, and YES to the rest). I am a little worried that some
unexpected relationships arise when better taxonomic sampling and better
phylogenetic data but, for the time being, they all seem to make sense.”
Comments from Claramunt: “I largely agree with Oscar’s assessment
and recommendations except for a few points. There is nothing haphazard in
Černý & Natale dataset: it is a supermatrix approach (not a supertree
approach) based on all GenBank sequences. Many species indeed lack data for
many loci but the methods they used are robust to missing data. The main
problem is the low statistical support for many clades.
“A. YES to rearrange
species in the genus Pluvialis
“B. YES to transfer wilsonia, collaris,
alticola, falklandicus, and nivosus to Ochthodromus. In this one, I disagree with the
recommendation, which is based on a conservative approach due to low support
for the relevant nodes. My point is that despite low support, the published
evidence points to the separation of this clade into three groups: Eupoda,
Anarhynchus, and Ochthodromus—and that classification is better,
in my opinion, than one with an Anarhynchus including most species.
First, is the issue of the meaning of the name, mentioned by Oscar; if
possible, we want names that make sense; there is nothing awkward with the beak
of all Ochthodromus species. But an additional argument is the
minimization of drastic changes in the meaning of the genera. Anarhynchus
was a monotypic genus restricted to New Zealand. Proposal C would transform it
into a species-rich cosmopolitan genus. This can be avoided by resurrecting Ochthodromus.
Anarhynchus may contain more than one species but will still be endemic
to Australasia.
“C. NO to Transfer Charadrius mongolus,
wilsonia, collaris, alticola, falklandicus, and nivosus to
Anarhynchus, for the reasons explained above.
“D. YES to adopt the
new linear sequence
“E. YES to transfer V.
cayanus to Hoploxypterus
“F. YES to transfer C.
modestus to Zonibyx
“G. NO to transfer C.
modestus to Phegornis. As much as I am averse to monotypic genera, modestus
and mitchellii are very divergent temporally and phenotypically.
“H. YES to recognize
the subfamilies Pluvialinae and Charadriinae.”
Comments from Stiles: “Given that the
clades of small “ex Charadrius” represent a massive polytomy, of the two
competing options (B and C) I agree with Santiago that B better expresses the
nomenclatural situation: the generic name Ochthodromus fits as a widely
distributed and relatively homogeneous group (and its English name also fits
nicely; whereas Anarhynchus was named for a definite oddball bill
of its type, and its two congeners are also with restricted Australasian
distributions. I therefore favor B, for separating mongolus to Eupoda,
maintaining a broad Ochthodromus, which in turn would produce a narrow,
separate Anarhynchus. Conversely, a NO to C (subsuming Ochthodromus into
Anarhynchus), which clashes with my preconceived notion of what a genus should represent. To sum up my votes on
this one, NO to C and G, Yes to all the other propositions."
Additional comments from Oscar Johnson: “Since
the writing of SACC proposal #979, an additional paper has been published on
the phylogenetics of the Charadriiformes that provides additional resolution,
in particular, to part of the tree relevant to the genus limits of the
Charadriidae. Dufour et al. (2024) used much of the same GenBank DNA data as in
Černý and Natale (2022), but with improved sequence alignment and quality
control methods that improved the final tree. Below I have included the
relevant portion of the tree of Dufour et al. (2024) showing the phylogeny of
the Charadriidae. This tree is time calibrated based on fossil data, and the
vertical axis marks are in 5 Ma increments.
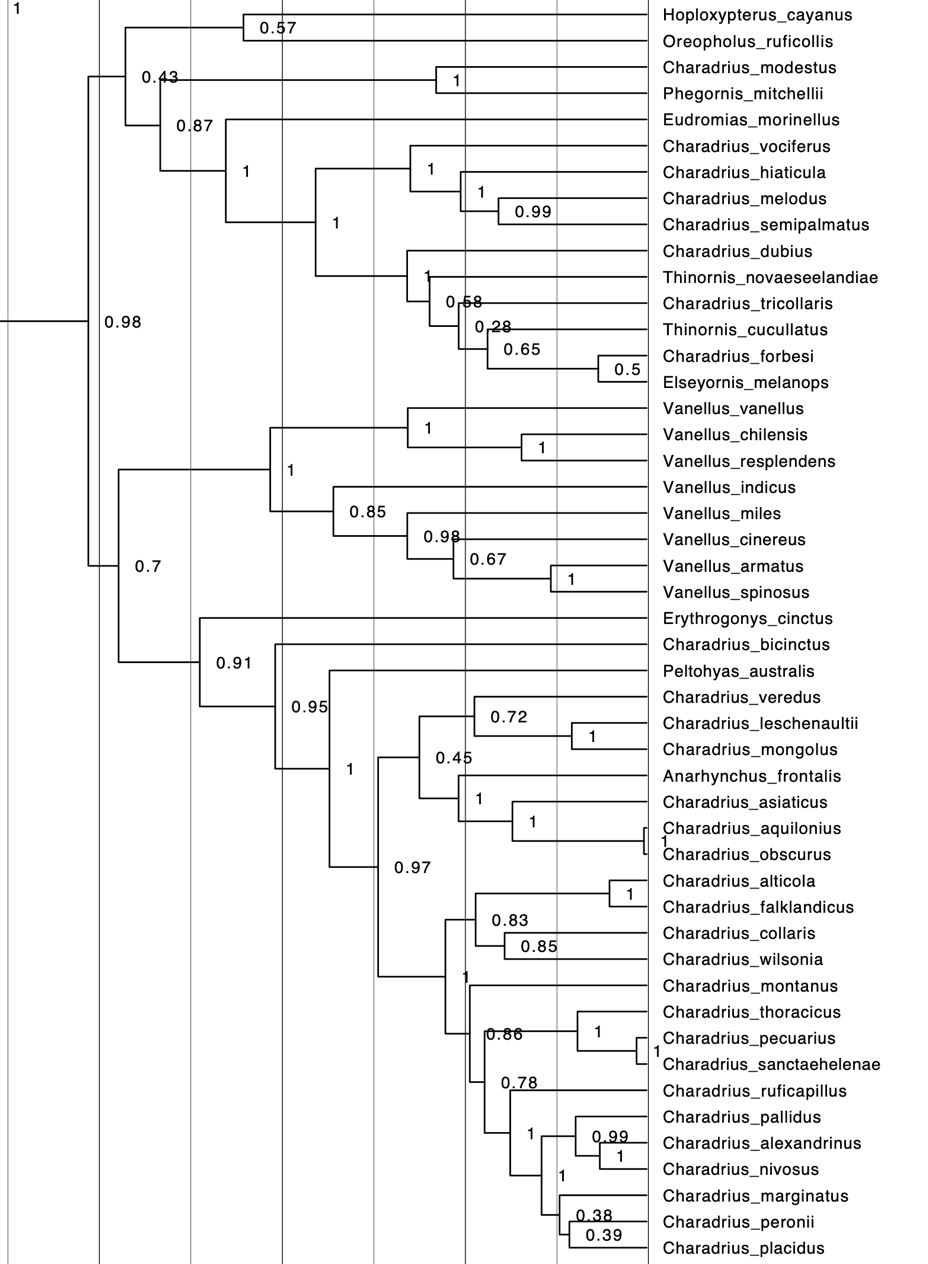
“Most
notable from this phylogeny is the confirmation (based on this set of DNA
markers) of the monophyly of the ‘Ochthodromus’ clade being separate
from the former Charadrius (=Anarhynchus). This Ochthodromus clade
had a 0.97 posterior probability, and in light of this (and comments by two
SACC members on the original proposal), I now recommend that this clade be
transferred to Ochthodromus instead of Anarhynchus.
“Unfortunately,
the resolution of “Eupoda” versus Anarhynchus still has very poor
node support (0.45 posterior probability) and should be considered a polytomy.
In light of this, I still recommend that the “Eupoda” Sand-Plovers be
transferred to Anarhynchus instead of their own genus.
“Many of
the other changes recommended in the original proposal are upheld by this
updated dataset, although cayanus and ruficollis are still on
long branches with poor support. The division of Thinornis from the
remaining Charadrius is also supported but all the species that would be
in Thinornis are extralimital.
“Dufour, P., P.-A. Crochet, F. L.
Condamine, and S. Lavergne. 2024. Seasonal migration and the evolution of an
inverse latitudinal diversity gradient in shorebirds. Global Ecology and
Biogeography e13817. DOI: 10.1111/geb.13817”
Comments
from Leo Joseph (voting for Del-Rio):
“A. Rearrange species in the genus Pluvialis: YES.
“B. Transfer Charadrius mongolus to Eupoda,
and wilsonia, collaris, alticola, falklandicus, and
nivosus to Ochthodromus: NO.
It's not that these shifts are totally unarguable and totally
indefensible, but one can counter, I think, that the molecular support for
robust structure, especially that for separating Eupoda, isn’t there.
True, it looks to be there in papers such as Barth et al. until you dig deeper.
Driving my caution here is that I have
learned the hard way that a clear mtDNA result can drive and override the
almost meaningless data from nuclear genes that often do little more than offer
a defence for saying, "Well, we haven't looked just at mtDNA." When I
dug into Barth et al.'s main paper and supplementary data, I saw that they had
one intron as the nuclear gene in their analyses and some very poor support for
structure in the whole group. So, despite what the mtDNA might be saying (and
remember they were focused on the position of one NZ taxon), I guess I'd prefer
to have the starting point of clear, robustly supported structure across
the group and then to recognize genera.
“C. Transfer Charadrius mongolus, wilsonia,
collaris, alticola, falklandicus, and nivosus to Anarhynchus:
YES. For all the same reasons as just outlined in suggesting NO, to Point B.
And yes it is a laugh that the oldest genus-group name should belong to an
obscure little NZ species with a weird bill, but it seems that that is the
case. In some ways, that might be a
telling point here because the entire group looks like a radiation of very
closely related things with some interesting evolution in bill morphology and
body size perhaps reflecting dietary/niche partitioning out on the world's
mudflats, coupled with an evolutionary shuffling and reshuffling of a limited
number of ancestral plumage traits (coloured chest bands and frontal bands for
example). Beware the nomenclaturalists of
the world who may find an older name than Anarhynchus out there. It does seem weird that a New Zealand bird
should have an older generic name than all of the others but maybe not if they
were all just unquestioningly placed in Charadrius.
“D. Adopt the new linear sequence: YES
“E. Transfer Vanellus cayanus to Hoploxypterus: YES.
Crazy bird in every way! Has always made me think of Elseyornis on
Strontium 90.
“F. Transfer Charadrius modestus to Zonibyx: YES.
“G. Transfer Charadrius modestus to Phegornis.
NO!
“H. Recognize the subfamily
Pluvialinae for the species in Pluvialis, and the subfamily Charadriinae
for the remaining species in Charadriidae. YES.”